

要旨
- 明治大学農学部生命科学科の田中博和准教授、同大学院農学研究科の荻田航佑(博士後期課程3年)、深澤成実(博士前期課程修了生)、王瑞安(博士前期課程1年)、藤崎翼(農学部卒業生)、片桐香(農学部卒業生)らの研究グループは、お茶の水女子大学の植村知博教授との共同研究により、アブラナ科植物のシロイヌナズナにおいて、SNARE タンパク質 SYP4 が PIN ファミリーのオーキシン注1輸送体の細胞膜における極性局在に重要であり、オーキシンに依存したさまざまな発生過程にも重要であることを明らかにしました。
- 本成果は、国際学術誌 Development に2025年10月22日に掲載されました。
概要
植物ホルモンの輸送体である PIN タンパク質注2は、細胞膜の一部の領域に偏って分布し、オーキシンを一方向的に排出することで極性輸送に重要な役割を担っています。これまでに、PIN タンパク質の局在制御において、トランスゴルジ網(TGN)に局在する ARF GEF BEN1 や Sec1/Munc18 タンパク質 BEN2/VPS45 などの膜交通因子が重要であることを当研究グループは明らかにしていました。BEN2/VPS45 は SNARE タンパク質を制御する種類の分子であると考えられていましたが、BEN2/VPS45 と共同で働き PIN タンパク質の局在を制御する SNARE タンパク質は同定されていませんでした。
本研究では、BEN2/VPS45 と SYP4 グループの SNARE タンパク質が互いを制御し、PIN タンパク質の極性局在の制御に関与することを明らかにしました。また、BEN2/VPS45 と SYP4 は、根の分裂領域の維持や子葉の維管束ネットワーク形成などのオーキシン依存的な発生過程に極めて重要であることも明らかになりました。
本研究は、JSPS 科研費(16H04806、22H02643)、JST CREST(JPMJCR20E5)の助成を受けて実施されました。
研究の背景
植物ホルモンの一種であるオーキシンは、PIN タンパク質などの排出輸送体により細胞の中から外に汲み出され、AUX1 などの流入輸送体により細胞の中に取り込まれることがわかっています。PIN1 や PIN2 などのタンパク質は、細胞膜の一部の領域に偏って分布することで、細胞の特定の方向にオーキシンを汲み出し、組織の中でのオーキシンの極性輸送に貢献していると考えられています。これまでの研究により、PIN タンパク質は細胞膜に安定に存在しているのではなく、エンドサイトーシスとエキソサイトーシス(細胞膜へのリサイクリング)を繰り返すことで細胞膜での偏りを保ったり、状況に応じて異なる細胞膜領域に移動すると考えられていました(図1)。田中准教授らはこれまでに、蛍光イメージングを活用したスクリーニングにより PIN タンパク質の細胞内での小胞輸送注3が異常になったシロイヌナズナの変異体 ben1 と ben2 を単離していました。また、ben1 変異体の異常の原因となっている遺伝子は低分子 G タンパク質の活性制御を担う ARF GEF の1つである MIN7/BIG5 をコードしており、ben2 変異体の原因遺伝子は Sec1/Munc18 ファミリーのタンパク質の一種である VPS45 をコードすることを見出していました。さらに、VPS45 はトランスゴルジ網(TGN)注4に局在する SYP4 グループの SNARE タンパク質と相互作用すること、ben2 変異によりその相互作用が弱まることも明らかにしていました。BEN1/MIN7/BIG5 と BEN2/VPS45 はどちらも TGN に分布する膜交通因子であることや、変異体の細胞レベルの表現型などから、BEN1/MIN7/BIG5 は TGN からの小胞の出芽に、BEN2/VPS45 は TGN への膜の融合に関わることで、PIN タンパク質の細胞内輸送に関わっていると推測されていました。また、BEN2/VPS45 は SYP4 SNARE タンパク質の制御を介して働くことが示唆されていました。田中准教授らのグループはben1;ben2 二重変異体は主根が短く、子葉の維管束が不連続になるなどのオーキシンに関連する異常を示すことが明らかにしていました。一方、
SYP4 ファミリーには3つのメンバー(SYP41、SYP42、SYP43)があり、植村教授らのグループは syp42;43 二重変異体や syp42;syp43;syp41/+ 個体では個体の成長が阻害されることや病原菌応答に異常が見られることを明らかにしていました。しかし、植物の発生の制御における VPS45 とSYP4 の関係性については明らかにされていませんでした。また、SYP4 が PIN タンパク質の局在制御やオーキシン依存的な発生制御においてどのような役割を担っているのかも不明でした。
また、野生型植物ではさまざまな組織において PIN タンパク質が細胞膜において非対称に局在していますが、根端の内皮組織における PIN1 タンパク質の分布と表皮組織における PIN2 タンパク質の分布に着目して詳しく解析したところ、ben1;ben2 二重変異体と syp42;43 二重変異体では野生型と比べて PIN タンパク質の偏在の度合いが低下していることも明らかになりました(図3)。syp42;43 二重変異体や ben2;syp42;43 三重変異体では根端の分裂領域が縮小したり、子葉の維管束が途切れたりと、オーキシンに関係していると考えられる発生過程に強い異常が生じていることもわかりました(図4)。これらの結果から、BEN2/VPS45 と SYP4 タンパク質は互いに局在を制御し、オーキシン輸送体の極性局在を通して、根の成長や子葉の維管束ネットワークの形成に貢献していると考えられます。
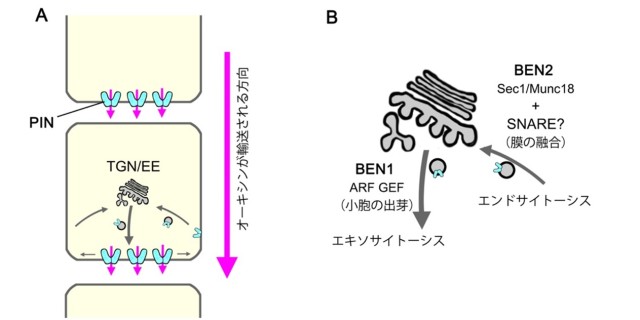
図1: 排出輸送体によるオーキシンの極性輸送のしくみと膜交通因子の働き
研究手法と成果
本研究では、BEN2/VPS45 が相互作用し、状態を制御していると推察されていた SNARE タンパク質 SYP4 に注目して、両者の関係性と SYP4 の発生制御における役割についての解析を行いました。まず、BEN2/VPS45 と SYP4 タンパク質の細胞内での分布を同時に可視化するために VPS45-RFP と GFP-SYP43 を同じ個体に発現させて、両者の分布を調べたところ、予想通りに局在部位が一致していることが示されました(図2)。次に、BEN2/VPS45 が SYP4 タンパク質を制御している可能性を調べるために、ben2 変異体における SYP4 タンパク質の局在の解析を行いました。その結果、通常は主に TGN に局在している3つの SNARE タンパク質が TGN に局在しにくくなりました(図2)。逆に、3つの SYP4 遺伝子のうちの2つの機能が損なわれた syp42;43 二重変異体では、VPS45 の TGN における局在がなくなりました。これらの結果から、BEN2/VPS45 と SYP4 SNARE タンパク質は、互いにTGNへの局在を促進していると考えられます。
また、野生型植物ではさまざまな組織において PIN タンパク質が細胞膜において非対称に局在していますが、根端の内皮組織における PIN1 タンパク質の分布と表皮組織における PIN2 タンパク質の分布に着目して詳しく解析したところ、ben1;ben2 二重変異体と syp42;43 二重変異体では野生型と比べて PIN タンパク質の偏在の度合いが低下していることも明らかになりました(図3)。syp42;43 二重変異体や ben2;syp42;43 三重変異体では根端の分裂領域が縮小したり、子葉の維管束が途切れたりと、オーキシンに関係していると考えられる発生過程に強い異常が生じていることもわかりました(図4)。これらの結果から、BEN2/VPS45 と SYP4 タンパク質は互いに局在を制御し、オーキシン輸送体の極性局在を通して、根の成長や子葉の維管束ネットワークの形成に貢献していると考えられます。
今後の展望
本研究で作出した変異体を詳しく解析すると、ben1; ben2 二重変異体と syp42; 43 二重変異体はオーキシン合成やオーキシン輸送の阻害剤に対する応答はどちらも異常を示すものの、それぞれに固有の特徴があることもわかりました。これらの違いは、BEN1/MIN7/BIG5 と BEN2/VPS45 が小胞の出芽と膜の融合という、異なる過程に関わっていることによるのかもしれません。今回の研究では BEN2/VPS45 の機能が弱まった変異体を用いて解析を行いました。VPS45 の機能が完全になくなった変異体は半数体の世代で致死となるため、VPS45 の機能がなくなった個体は得られていませんが、今後、VPS45 の機能がさらに弱まった植物を作り出すことができれば、それぞれの因子の発生における役割をより詳しく理解することができると期待されます。また、ben2; syp42; 43 三重変異体では胚軸が太くなるという、不思議な現象も観察されています。オーキシンの極性輸送は胚軸の縦方向の伸長や、それと関連した現象である屈性に関わることはよく知られていますが、太さの制御に関わっているかは不明です。三重変異体の胚軸が太るしくみをさらに調べていくことで、小胞輸送系因子の役割についての新しい知見が得られる可能性があります。
また、3つの SYP4 は発生過程の制御における貢献度が異なることも見出されています。それぞれの因子が異なる機能を持っている場面もあるかもしれません。今後、個別の分子の違いに注目して詳しい解析を行うことで、膜交通因子の役割と機能分化についての理解が深まると期待されます。
用語説明
- 注1 オーキシン
オーキシンは細胞伸長や器官形成、細胞分裂の制御などの様々な現象に関わる植物ホルモンであり、植物が光に向かって成長することを研究していたダーウインが「動くシグナル」として記載したことでも有名です。インドール-3-酢酸などの化合物がオーキシンの実体です。
- 注2 PIN タンパク質
pin-formed1 (pin1)という、花芽が形成されずに花茎が針のようになってしまうシロイヌナズナの変異体の原因の遺伝子として PIN1 遺伝子が発見されました。PIN1 タンパク質は細胞膜の一部の領域に局在し、細胞の中から外にオーキシンを排出することで組織におけるオーキシンの極性輸送に重要な働きをしています。シロイヌナズナでは PIN1 とアミノ酸配列が良く似たタンパク質で、細胞膜に局在している PIN2、PIN3、PIN4、PIN7 などのタンパク質もオーキシンの輸送に働いています。
- 注3 小胞輸送
タンパク質や脂質などの物質がオルガネラや細胞膜などの間を移動する際に膜小胞を介して輸送されることを小胞輸送と呼びます。メンブレントラフィックや膜交通とも呼ばれます。この過程では小胞を作り出すためのタンパク質や、小胞を標的の区画に結合させるための「膜交通因子」が働きます。ARF GEF は小胞の出芽や積荷の選別に、Sec1/Munc18 タンパク質と SNARE タンパク質は膜の融合に関わるタンパク質です。
- 注4 トランスゴルジ網・初期エンドソーム
トランスゴルジ網(TGN)は、ゴルジ体のトランス側に存在する入り組んだ形をした区画で、細胞膜や液胞に運ばれる物質を選別して小胞に積み込まれる選別の場となるオルガネラです。リボソームで合成された膜タンパク質はゴルジ体の中で修飾され、TGN に移動して目的の区画に運ばれます。一方、エンドサイトーシスされた物質は最初に初期エンドソーム(EE)という区画に運ばれて選別されると考えられています。植物細胞においては、細胞膜からエンドサイトーシスで取り込まれたトレーサーはまず TGN に輸送されることから、初期エンドソームは TGN と同一の区画であると考えられています。
参考図
図1: 排出輸送体によるオーキシンの極性輸送のしくみと膜交通因子の働き
(A) PIN タンパク質によるオーキシンの極性輸送の模式図。PIN タンパク質(水色)が細胞膜の一部の領域に偏って分布し、オーキシンを排出することで、組織の中で一方向的にオーキシンが輸送される(マゼンタ矢印)。
(B) 膜交通因子 BEN1 と BEN2 の推定される働き。BEN1/MIN7/BIG5 は TGN に局在し、小胞の出芽に関与すると推察される。BEN2/VPS45 はトランスゴルジ網において膜の融合を担う SNARE タンパク質を制御すると考えられる。
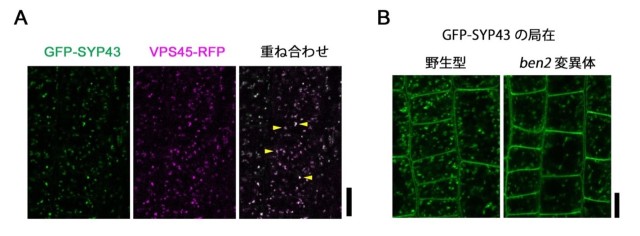
図2: BEN2/VPS45 は SYP4 と共局在し、SYP4 の TGN 局在を促進している
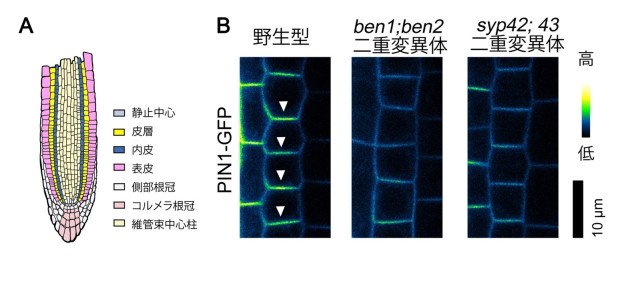
図3: ben1;ben2 二重変異体と syp42;43 二重変異体はPIN1の極性局在に異常を示す
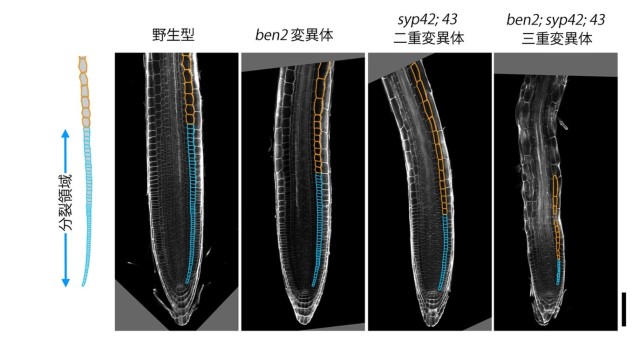
図4: BEN2/VPS45 とSYP4 は根の発生に重要である
図2: BEN2/VPS45 は SYP4 と共局在し、SYP4 の TGN 局在を促進している
(A) GFP-SYP43 と VPS45-RFP を同時に発現するシロイヌナズナの根端の表皮細胞の共焦点顕微鏡像。GFP-SYP43 (緑)と VPS45-RFP (マゼンタ)の蛍光シグナルを重ね合わせると細胞内の同じ場所(TGN)に分布していることがわかる(矢尻)。
(B) 野生型と ben2 変異体の根端の表皮細胞における GFP-SYP43 の局在。野生型においては斑点上の TGN 局在が目立って見えるが、ben2 変異体では細胞膜局在が増加している。スケールバーは10 µm を示す。
図3: ben1;ben2 二重変異体と syp42;43 二重変異体はPIN1の極性局在に異常を示す
(A) 根端の縦断面の模式図と組織の種類。
(B) 野生型と変異体の内皮組織における PIN1-GFP の分布のカラーコード表示。強いシグナルは黄色で、弱いシグナルは青色で表示している。野生型では内皮細胞の基部側(根端側)の細胞膜にPIN1-GFPの強いシグナルが見えるのに対して、ben1;ben2 二重変異体と syp42;43 二重変異体では偏在が弱まっている。スケールバーは10 µm を示す。
図4: BEN2/VPS45 とSYP4 は根の発生に重要である
野生型植物の根の先端には活発に細胞分裂を行う小さい細胞の集団(分裂領域)が見られる。3つの SYP4 遺伝子のうちの2つの機能を失った syp42; 43 二重変異体や、さらに VPS45 の機能も弱まった ben2; syp42; 43 三重変異体では分裂領域が狭まっている。スケールバーは 100 µm を示す。
論文情報
題目
VPS45 and SYP4 Qa-SNARE proteins jointly regulate auxin distribution and plant development in Arabidopsis
著者名
Kosuke Ogita, Narumi Fukasawa, Ruian Wang, Tsubasa Fujisaki, Kaori Katagiri, Tomohiro Uemura, Hirokazu Tanaka.
論文誌名
Development