





■要 旨
●今回の発見は、植物の光合成や成長に重要な働きをする気孔が、環境情報を細胞レベルで記憶していることを示すものであり、その仕組みの解明に繋がる分子メカニズムが明らかとなりました。
●本研究成果は、英国際誌 Scientific Reports において2019年7月22日19時(日本時間)に公開されました。
■概 要
■研究の背景
■研究手法と成果
さらに、このような長日条件に依存した気孔開口促進が、植物を短日条件に戻した場合でも維持されているかどうかを調べるために、長日条件で育てた植物を短日条件に移し、1週間後に気孔開度を調べたところ、短日条件に移した後も、少なくとも1週間は気孔の開口促進が維持されていることを発見しました(図3)。これと一致して、SOC1の発現も短日条件に移した後も、少なくとも1週間は高いレベルで維持されていました。一方で、SOC1の発現を誘導するFTの発現は、短日条件に移した後1週間では発現が低下していました。以上の結果から、植物の孔辺細胞は、少なくとも1週間はFTに依存せずに、気孔開口促進とSOC1の発現上昇を維持する記憶システムを持っていることが明らかとなりました(図3)。
この記憶システムのメカニズムを探るために、遺伝子発現に重要な役割を果たすことが知られているヒストンのメチル化とアセチル化の状態注7)を調べたところ、SOC1遺伝子周辺では、FT依存的にヒストンのメチル化が引き起こされていることが明らかとなり、ヒストンのメチル化(ヒストンH3K4)が孔辺細胞の記憶を担うメカニズムの一つである可能性が示唆されました(図4)。
■今後の期待
本研究で明らかとなったように、花成ホルモンFTは、日長に依存した気孔開度調節で重要な役割を果たしますが、日長だけではなく、温度もFTの発現に強く影響することが知られています(気温が高いと発現が高まり、気温が低いと発現が低下する)。よって、植物は、気温が変動しても安定的にFTの作用を持続させるための一つのメカニズムとして、ヒストンの修飾を介してSOC1の発現制御を行なっているのかもしれません。これは、変動する温度条件下で安定的に日長に応答するための植物の戦略機構の一つである可能性が考えられ、さらなるメカニズムの解明をする必要があります。
また、気孔は光合成に必要な二酸化炭素を植物が取り入れるための唯一の入り口であり、その気孔が大きく開口することは光合成の促進につながります。シロイヌナズナにおいて、長日条件下では、花芽形成とそれに続く種子形成・成熟に多くのエネルギーが必要となります。よって、植物は盛んに光合成をする必要があり、この期間に気孔を大きく開かせることは植物の生育と子孫繁栄に重要な応答なのではないかと私たちは考えています。
用語説明
アブシシン酸(ABA)は、植物ホルモンの一種で、乾燥などのストレスに対応して合成される。気孔の閉鎖や種子の休眠、生長抑制などを誘導する。
注2)細胞膜プロトンポンプ
ATPをエネルギーとして、細胞の内側から外側に水素イオンを輸送する一次輸送体。細胞膜を介して形成される水素イオンの濃度勾配は、さまざまな物質を輸送する二次輸送体の駆動力として利用されています。気孔孔辺細胞においては、青色光により活性化され、カリウム取り込みの駆動力を形成し、気孔開口を引き起こすことが知られています。
注3)モデル植物シロイヌナズナ
アブラナ科の一年草で、ゲノムサイズが小さく、世代期間が短く、室内での栽培が可能で、形質転換が容易などモデル生物としての利点を多く備えているため、植物のモデル生物として盛んに研究に用いられている。2000年に植物として初めて全ゲノム解読が終了した。
注4)光周性花成誘導に関わる因子(花成ホルモンFTや転写因子SOC1など)
光周性とは生物が日長の季節変化を認識し、それぞれの種にとって1年の中で最適と考えられるタイミングで花芽形成のような生命活動を行う性質のことをいう。これに関わる因子として、花成ホルモン「フロリゲン」の実体であるFTやその下流因子で転写因子のSOC1が知られており、これらが働くことにより、日長に応じた花芽形成が誘導されることが明らかとなっている。
注5)気孔コンダクタンス
気孔抵抗(大気と葉内の気体のやりとりの際、気孔において生じる拡散抵抗)の逆数であり、気孔を通した気体(特に水蒸気とCO2)の通りやすさを表す。気孔が大きく開口しているほど気孔抵抗は小さくなり、気孔コンダクタンスは大きくなる。逆に、気孔が閉じていると気孔抵抗は大きく、気孔コンダクタンスは小さくなる。
光合成が起こるためには、CO2が大気から葉内へ、そして葉肉細胞の葉緑体へ拡散し、最終的にルビスコまで到達する必要がある。その拡散速度は、拡散経路における抵抗に依存しているが、CO2拡散は主に気孔抵抗により制限されることから、気孔コンダクタンスは光合成活性に大きく影響している。
注6)孔辺細胞プロトプラスト
葉の表皮組織を単離し、酵素処理によって細胞壁を消化した細胞(プロトプラスト)をサイズによってより分けて、気孔の孔辺細胞だけを集めたもの。
注7)ヒストンのアセチル化とメチル化
核内において遺伝子情報がコードされているDNAはヒストンと呼ばれる8量体のタンパク質に巻き付いて存在している。遺伝子の発現制御にヒストンテールと呼ばれるN末端・C末端領域のアミノ酸の修飾状態が関係していることが知られている。今回の研究ではヒストン3(H3)のアミノ酸のN末端から4番目のリシン(K4)のトリメチル化(me3)とK9のアセチル化(Ace)をそれぞれ認識する抗体を用いてヒストンテールの修飾状態を調べた。この2つの修飾はいずれも、遺伝子発現に対して促進的に作用することが知られている。
参考図
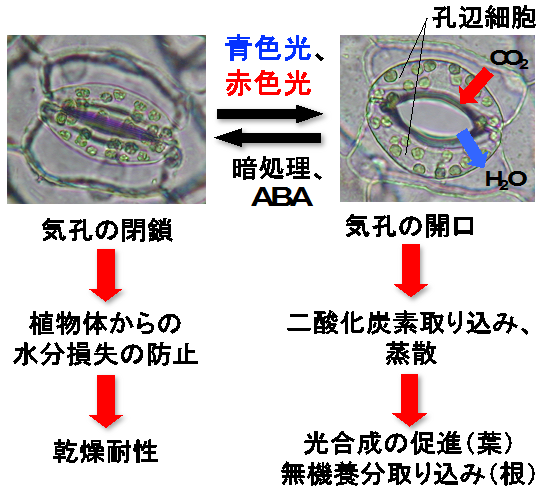
図1 気孔の開閉とその働き
気孔は青色光や赤色光によって開口し、暗処理や乾燥ストレスにより生合成される植物ホルモン・アブシシン酸(ABA)により閉鎖します。気孔は、光合成に必要な二酸化炭素の唯一の取り込み口として働きます。
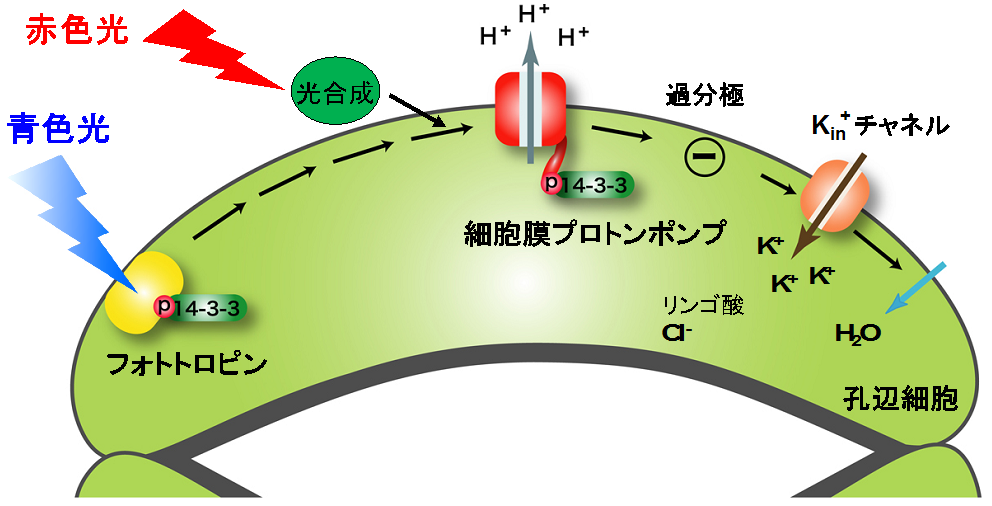
図2 光による気孔開口
青色光は、青色光受容体フォトトロピンに受容され、細胞膜プロトンポンプを活性化し、カリウムイオン(K+)取り込みを誘導します。これにより、浸透圧が上昇し、水が取り込まれ、孔辺細胞の体積が増加することで気孔が開口します。赤色光は、葉肉細胞や孔辺細胞の葉緑体における光合成を介して細胞膜プロトンポンプを活性化し、気孔開口を誘導します。
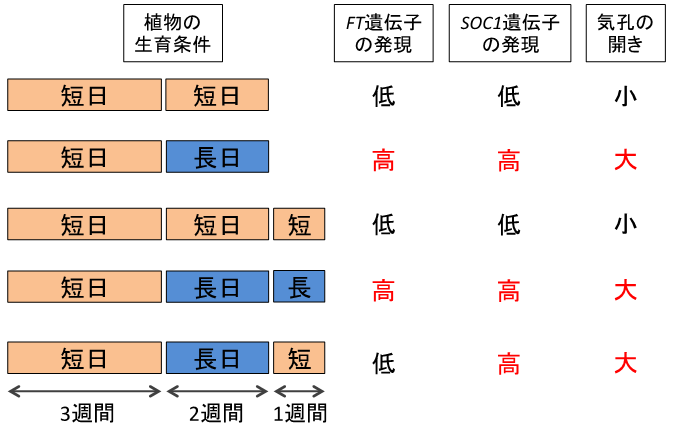
図3 日長に応じた気孔の開きと孔辺細胞における遺伝子発現
長日条件(16時間明期)で育てた植物は、短日条件(8時間明期)で育てた植物に比べて気孔が開きやすくなっており、このとき孔辺細胞内ではFT遺伝子やSOC1遺伝子の発現が高まっていた。
長日条件で誘導された気孔開口促進は、短日条件に移してから1週間経っても維持されており、FTの発現は低下していたが、SOC1の発現は維持されていた。
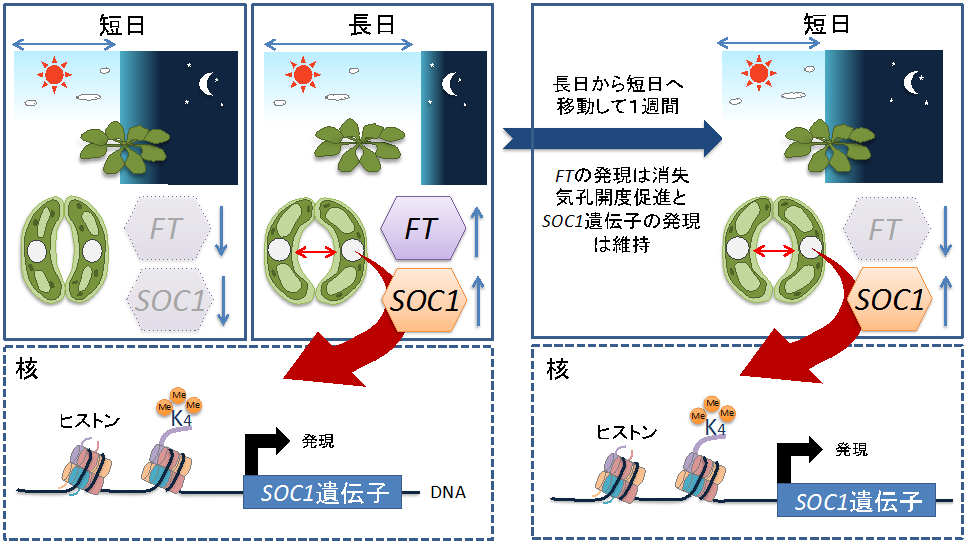
図4 気孔孔辺細胞における日長の記憶
短日条件と比較すると長日条件では、光による気孔開口が増加する。孔辺細胞においても長日条件では光周性花成誘導にかかわるFT遺伝子の発現が上昇する。すると、転写因子SOC1遺伝子の発現が誘導され、同時に、FT依存的にSOC1遺伝子近傍のヒストンH3K4のトリメチル化が引き起こされる。さらに、長日から短日に移し、1週間は気孔の開口促進が維持され、その際、FTの発現は減少したのに対して、SOC1の発現は高いレベルで維持される。植物の孔辺細胞は、少なくとも1週間は、FTに依存した気孔開口促進とSOC1の発現上昇を維持する記憶システムを持っていることが明らかとなり、ヒストンH3K4のトリメチル化が記憶のメカニズムである可能性が示唆された。
論文名:“Regulation of stomatal opening and histone modification by photoperiod in Arabidopsis thaliana.” (日長による気孔開口とヒストン修飾の調節)
著者:Saya Aoki*, Shigeo Toh*, Norihito Nakamichi, Yuki Hayashi, Yin Wang, Takamasa Suzuki, Hiroyuki Tsuji, Toshinori Kinoshita(*共筆頭著者)
DOI:https://doi.org/10.1038/s41598-019-46440-0
論文公開:2019年7月22日午後7時(日本時間)/ 2019年7月22日午前10時(英国GMT時間)
※本件につきまして、取材をご希望の方はこちらのフォームからご連絡ください。