

岐阜大学応用生物科学部の修士二年生羽賀夏子さん、同学部山根京子准教授、静岡県農林技術研究所伊豆農業研究センターわさび生産技術科馬場富二夫科長、明治大学農学部矢野健太郎教授、大鐘潤教授らは、日本のワサビ属植物のルーツを、葉緑体全ゲノム解読から明らかにしました。
本研究成果は,英国の国際誌「Scientific Reports」(電子版)2019年10月7日付(日本時間午後6時)に掲載されました。
【発表のポイント】
・ 日本のワサビ属植物2種7系統の葉緑体全ゲノムを世界で初めて解読した
・ 日本固有種であると考えられていたユリワサビの中には、大陸のワサビ属植物に近縁な系統が含まれていた
・ 日本固有と考えられるワサビ属植物系統群は約130万年前に分岐したことがわかった
・ 栽培ワサビのルーツに北方系野生ワサビはおそらくは関与していない
・ 栽培ワサビとなった野生ワサビは日本固有種であり、氷河期時代に日本海側気候に適応した日本海要素植物であることがわかった
・ 葉緑体全ゲノム配列約15万塩基対の比較解析から、日本の主要なワサビ3品種を区別するDNAマーカーの構築に成功した。これらは品種の特定に用いることができ、品種改良や保全活動への応用が期待できる
【概要】
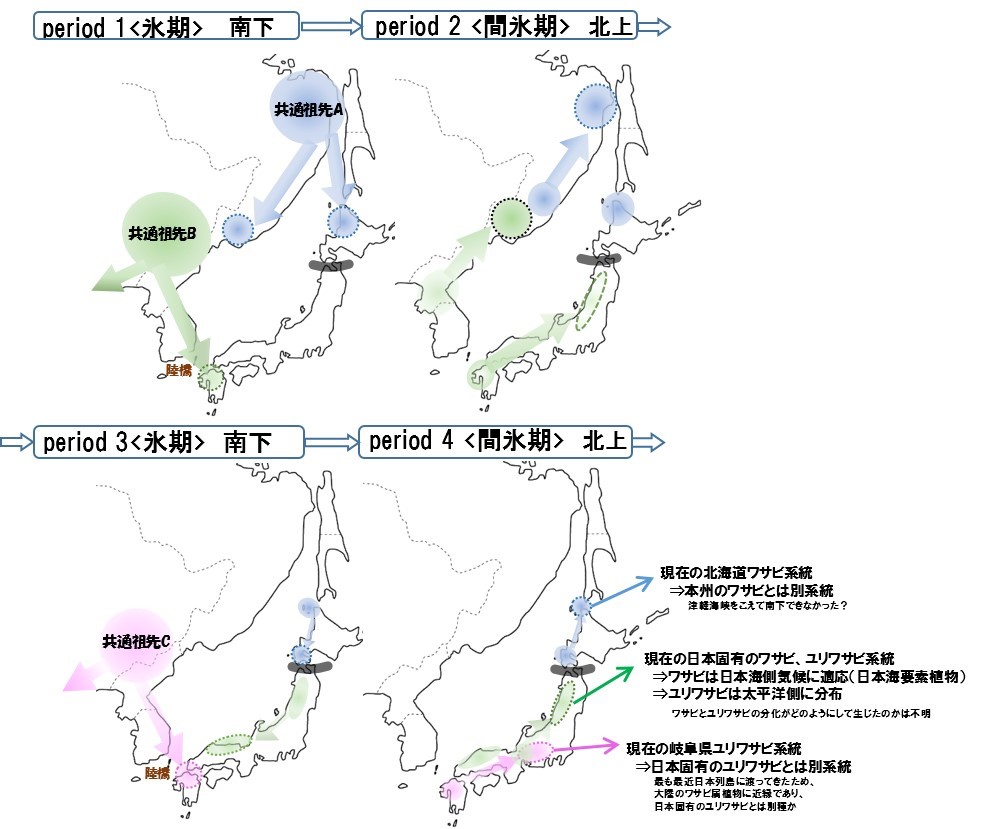
日本にはワサビとユリワサビという、二種類のワサビ属植物が生育しています(写真1)。日本人が薬味として利用してきた栽培ワサビは、野生のワサビが栽培化されたものです。ところが野生ワサビのルーツに関しては全くわかっていませんでした。「日本のワサビ属植物は、もともと日本列島の形成時からいた植物なのか?」あるいは「大陸からわたってきた植物なのか?」—それすら不明でした。そこで今回、葉緑体の全ゲノム配列を用いて、大陸の近縁なワサビ属植物との比較解析を行い、日本のワサビ属植物がどのように大陸から移動し、分布を広げたのかを調べました。その結果、岐阜県に自生するユリワサビが、他の日本のユリワサビよりも、日本から遠く離れた中国雲南省に自生するワサビ属植物と遺伝的に近縁であることがわかったのです。ユリワサビと考えられていた一部の系統は、日本固有のユリワサビとは異なる種である可能性が考えられます。また、分岐年代の推定から、この二種間は比較的最近(48万年前)に分岐したことがわかりました。つまり、この岐阜県のユリワサビは、他のユリワサビとは別の時代に、別の祖先系統から日本にわたってきたと考えられます。その他の日本のワサビ属植物は、分岐年代推定の結果、少なくとも約130万年前に大陸のワサビ属植物とは分化したことが明らかになりました。ただし、この日本のワサビ属植物のうち北海道のワサビは、本州および九州のワサビ属植物とは遺伝的な分化の程度が大きかったことから、少なくとも現在のワサビ品種は、北方系のワサビ属植物はルーツでないと考えられます。
以上の結果を図にまとめたものが図1になります。日本のワサビ属植物は、第四紀の氷河期時代の氷期に、陸続きになった際に大陸からわたってきた祖先に由来し、しかもこうした大陸からの移動は複数回あった可能性が高いこともわかりました。現在、野生ワサビは主に日本海側の多雪地帯に分布しています。野生ワサビの祖先種が生育していた大陸とは環境が大きく異なっていたことが予想されることから、野生ワサビは、氷河期時代に列島に渡ってきた後、日本の多雪地帯に適応し、ブナなどに代表される「日本海要素植物」として成立したと考えられます。つまり、野生ワサビは、氷河期時代に日本の環境に適応して成立した日本固有種であることが、あらためて示された結果となりました。
本研究ではさらに、葉緑体全ゲノム配列を用いて主要3品種を区別するDNAマーカーを構築しました。本マーカーを用いれば、品種改良や保全集団の選定など、幅広い分野への活用が期待できます。
【研究内容】
ワサビは学名をEutrema japonicum (Miq.) Koidz. とし、日本固有種と考えられてきました。日本にはワサビとは別に、ユリワサビ[E. tenue (Miq.) Makino]という野生植物も自生しています。ユリワサビにはワサビのような立派な根茎は無く、ゆり根のような鱗片状根茎ができます(写真1)。二種は、それぞれ典型的な形態をしている個体どうしを見比べると、植物体の大きさも、花の大きさ、形態、根茎の形など、かなり違いがみられるため、両者の区別は簡単です(写真2)。また、生育している場所も、ワサビに比べてユリワサビは標高が低く、水際というよりは渓流沿いの湿った斜面に自生するなどの違いがあります。
 ②
②




⑥

一方、ワサビ属植物[Eutrema]は主に東アジアに分布するアブラナ科植物で、26種知られています。本属で栽培化された植物はワサビだけです。このうち日本のワサビ属植物に形態学的にも最も近い植物がシャンユサイ[E. yunnanense]とよばれる野生種で、主に中国雲南省に自生しています。
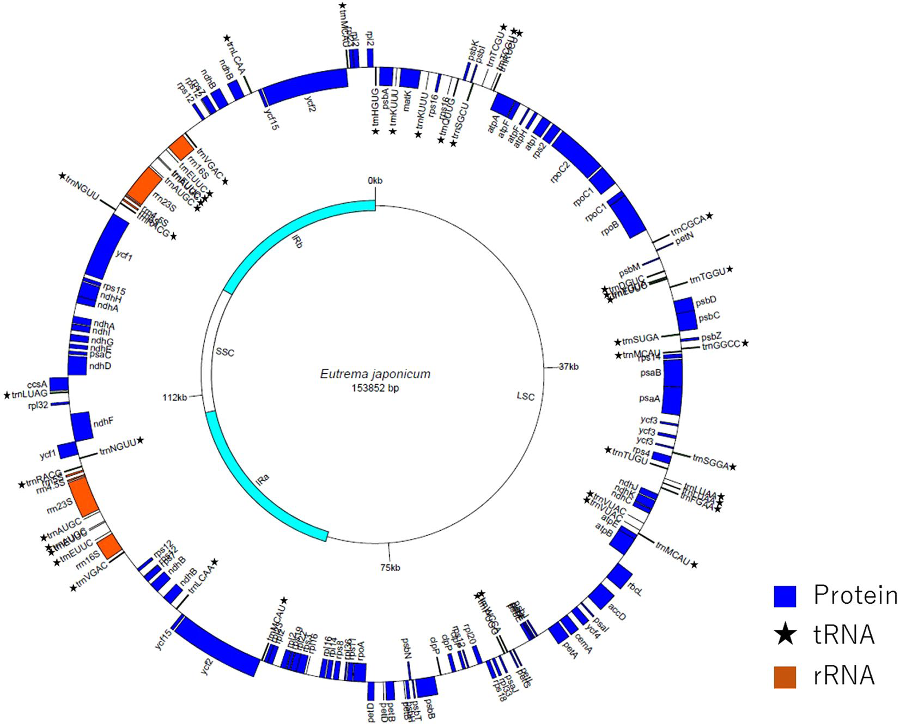
ワサビは、和食に欠かせない食材として日本における重要な栽培植物ですが、遺伝や進化の研究は行われてきませんでした。近年は「ゲノム」とよばれる生命の維持に必要な最小限の単位を意味する遺伝情報が様々な生物で解読されニュースになっています。ワサビでは、葉緑体とよばれる細胞小器官に存在する小さなゲノムですら解読されていませんでした。そこで本研究では、次世代シークエンサーとサンガー法を用いて7系統の配列を解読しました(図2)。
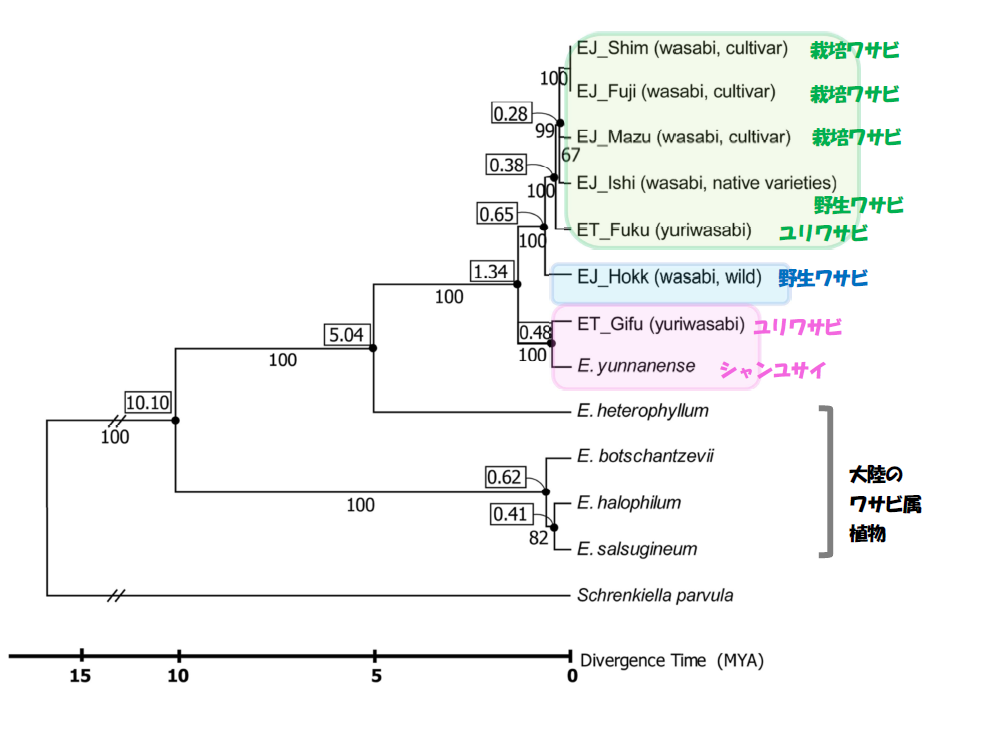
葉緑体ゲノムの塩基配列は、倍数化や組換えの影響を受けないため、進化の系譜を考察するために植物では有用なツールとして汎用されています。今回得られた葉緑体全ゲノム配列のうち、遺伝子がコードされている79全ての領域の配列を用いて、すでに解読されている大陸の近縁なワサビ属植物5系統と一緒に比較解析を行いました(図3)。その結果から、以下のことが示されました。
(1) 岐阜県のユリワサビが中国雲南省の系統(シャンユサイ)とグループを形成した。この二種間の分岐は48万年と比較的最近であった
(2) 岐阜県のユリワサビとシャンユサイのグループと、その他の日本のワサビ属植物は、約130万年前に分岐した
(3) 北海道のワサビは、他の本州のワサビ属植物からは、分岐系統学的にはなれた場所に位置した
(4) 主要栽培3品種は同一グループを形成した
(1)と(2)に関しては全くの予想外で大きな問題を含んでいました。詳しく説明しましょう。図3をみると、ユリワサビとワサビは単一グループを形成しなかった(=多系統)ことから、これからはそれぞれ独立した種とみなせない可能性があります。山根は、最近の複数の系統を用いた解析や現地情報、交配実験のデータからも、ワサビとユリワサビは、独立の種としては認めず「ワサビーユリワサビ複合種」とした方がよいとの考えに至っています。仮に、複合種を認めても、認めなかったとしても、ユリワサビのうちの1系統が、大陸のシャンユサイとグループを形成したことは大きな問題となります。この系統樹に従うと、日本固有と考えられていた種が、「側系統」となります。「側系統」とは分岐系統学で用いられる専門用語ですが、今回のような樹型が得られた場合は、日本のユリワサビもワサビも、独立の種としては認めずに、E. yunnanenseのなかに含めなければならなくなるのです。つまり、日本のワサビ属植物の固有性は否定されることになります。この点は論文でも慎重に議論しましたが、近年、大陸でもE. tenueと分類された集団が複数発見されていることから、岐阜県のユリワサビの系統は、他のユリワサビとは別種であるとすることで、残りのユリワサビとワサビの系統群(図1で示した緑色のグループ)の日本固有性が支持されました。
さらに、この結果は、日本列島に、大陸から異なるタイミングで渡ってきたワサビ属植物の存在を示すことになり、「氷河期時代に陸続きになったこと」、「その際に植物が移動したこと」が、推定した分岐年代からも裏付けられたともいえます。興味深い結果といえるでしょう。
(3)に関しても、今回の結果は予想外でした。ワサビは標高の高い冷涼な環境に好んで生育しているため、北方系の植物の要素を持つことが予想されていました。ところが今回の結果は、その可能性は低いことを示していました。つまり、(3)で示した栽培ワサビ系統は、北海道の野生ワサビよりむしろ、九州のユリワサビの方が、系統学的に近かったのです。しかも、北海道の野生ワサビは、本州と九州の「ワサビーユリワサビ系統群」からは系統樹でも離れて位置しており、これらは、図1で示したとおり、他のグループとはルーツを別にしている可能性が示されました。この結果から、日本の栽培ワサビの祖先的な野生集団は、北方ルートで渡ってきたワサビ属植物ではなく、図1で示したようなルート(緑色)を起源とすることが示唆されたのです。この、北海道の野生ワサビに関しては、現在のところ形態上の明確な違いは見つかっていませんが、樺太地方に自生する「カラフトワサビ」である可能性があり、さらに詳しく調べる必要があります。
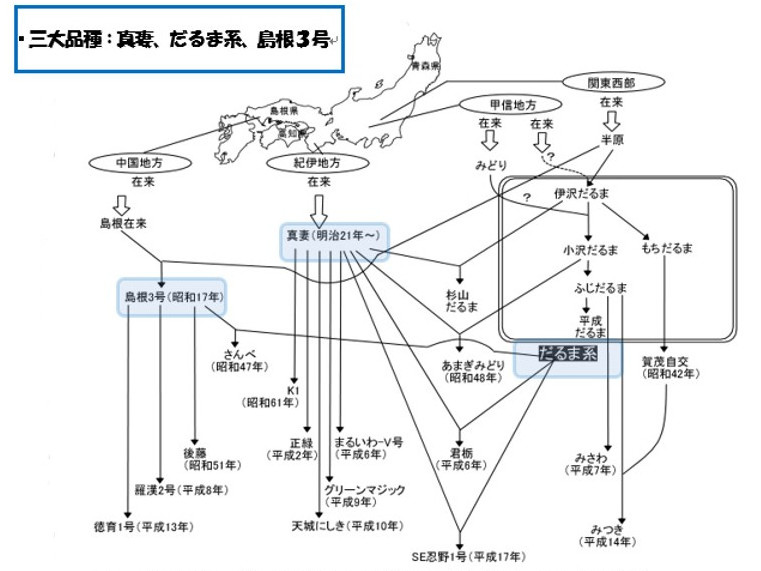
(4)に関連して最初に、ワサビの品種について説明しましょう。あまり知られていないかもしれませんが、ワサビにはたくさんの品種が存在します。農林水産省の品種登録サイトにも約20品種が登録されています。これまで知られている主要な品種の来歴を、文献や聞き取り調査から調べた系譜を示します(図4)。たくさんの品種が作出されてきたなかで、品種改良の母本となる基本的な系統は、主要な3系統(’だるま系’、’島根3号’、’真妻’)であることがわかります。
こうしたなか、品種を判別するためのDNAマーカーづくりに際し、予備的な実験を行ったところ、どうしても‘ふじだるま’と‘島根3号’を区別する多型サイトを見つけることができませんでした。つまり、両品種の母系統は遺伝的にかなり近く、区別するマーカーが見つけられなかったのです。そこで、葉緑体の全ゲノム配列を解読することで、両品種間の違いを見つけようと試みました。その結果、15万塩基対のなかで、両品種を区別するサイトは、たった2サイトしかないことがわかったのです。そこで今回、この2サイトのうちの1つを用いてDNAマーカーを構築し、両品種を区別することが可能になりました(図5)。葉緑体の全ての塩基配列を解読して初めて可能になった識別でした。
今回構築した品種判別DNAマーカーは、あくまでも母系マーカーであり、花粉親の系統は特定できませんが、その後の多数の品種や系統を用いた結果から、現在市場に出回っている品種や、全国で商業目的に栽培されるワサビは、現在のところ例外なく今回明らかにした3タイプのDNA型しかみられないことがわかっています。このうち、’真妻’は毎年品評会で上位を占める優良品種です。’真妻’を他と区別できるマーカーが構築できたことで、品種の誤認などを回避することが可能になりました。また、’島根3号’は、’島根在来’と’半原’という病気に強い品種を自然交配により掛け合わせてできた品種であると言われてきましたが、’島根3号’の母系は、’半原’の可能性が高いことがわかりました。こうした情報を利用して、「’島根在来’さがし」の役立つツールとして用いることが可能になりました。その他「野生ワサビか栽培ワサビの逃げ出しか」の判定に、すでに今回構築したDNAマーカーは頻繁に利用されています。今後は、より詳細な品種判別を行うためにも、核DNAマーカーを開発し、より詳しく判別できるマーカーの開発が急務となっています。
【今後の展開】
本研究の目的は、大陸のワサビ属植物との比較解析を行うことで、日本のワサビ属植物のルーツを明らかにすることでした。現在は、日本全国から収集した約300系統のワサビ属植物の系統解析を行い、より詳細な分析を進めています。その結果、多くの系統を分析した結果も、今回提示した進化モデルとは矛盾がないことがわかってきました。たとえば、本研究では、中国のシャンユサイと系統学的に近い集団は岐阜県由来のユリワサビのみ供試されていましたが、その後の解析から、別の岐阜県集団や、山梨県、群馬県にも類似系統が存在することがわかったのです。ただ、これらが本当に、E. tenueとは別種であるのかどうかについては、詳細に調べる必要があります。
こうして得られた「ワサビの来た道」をふまえたうえで、現在は「日本のワサビはなぜ辛いのか」という疑問を解明するために研究を進めています。最近、海外の研究グループがワサビ属植物の辛味成分の前駆体物質を調べた論文を発表しました(Hao et al., 2016)。その結果、日本のワサビ属植物の辛味関連成分は大変ユニークであることがわかってきたのです。我々の研究室で現在も辛味関連成分を調べていますが、大変興味深いデータが得られています。
辛味関連成分が日本で独自に進化したとすると、今回の結果から、約100万年の間の第四期の氷河期時代に生じたと推測されます。日本列島に渡ってきたワサビ属植物が、氷河期時代にどのようにして独自の辛味成分を持つようになったのかを、豊富な材料を用いることで、分子レベルで明らかにしたいと考えています。
子供たちからはよく「ワサビはなぜ辛いの?」という質問を受けます。簡単な質問にみえて実は答えるのが大変難しく、本質的な問いなのだということが、研究をすすめるなかでみえてきました。現地調査や分子や化学の実験など、多角的な視野からこの本質的な問いに対する答えを見つけるべく、研究を進めています。
岐阜大学の山根研究室では、系統保存と保全活動も行っています。野生ワサビが自生する環境が年々変化し、集団の消失もみられます。将来のワサビ属植物の分布域をシミュレーション予測したところ、現在の野生ワサビの分布域は、地球温暖化により深刻な影響を受けることが示されました。日本においては、温暖化の影響として日本海側の多雪地帯の積雪量の減少が深刻になるという予測がされており、野生ワサビの自生地への影響が懸念されます。氷河期時代を生き残り、高い遺伝的多様性が維持されてきた日本のワサビ属植物は、遺伝資源として、未だ知られていない多くの可能性を秘めていることがわかってきています。日本が誇る貴重な資源を守るためにも、今後も活動を続けてゆきたいと考えています。
【論文情報】
雑誌名:Scientific Reports
タイトル:Complete chloroplast genome sequence and phylogenetic analysis of wasabi (Eutrema japonicum) and its relatives
著者:Natsuko Haga 1 , Masaaki Kobayash i2 , Nana Michiki 1 , Tomoyuki Takano 2 , Fujio Baba 3 ,
Keiko Kobayashi 1 , Hajime Ohyanagi 2,4, Jun Ohgane 2 , Kentaro Yano 2 & Kyoko Yamane1
1Gifu University, Faculty of Applied Biological Sciences, 1-1 Yanagido, Gifu City, Gifu, 501-1193, Japan. 2School of Agriculture, Meiji University, 1-1-1 Higashi-Mita, Tama-ku, Kawasaki, Kanagawa, 214-8571, Japan. 3Izu Agricultural Research Center, Shizuoka Prefectural Research Institute of Agriculture and Forestry, 3012 Inatori, Higashiizu-cho, Kamo, Shizuoka, 413-0411, Japan. 4King Abdullah University of Science and Technology (KAUST), Computational Bioscience Research Center (CBRC), Thuwal, 23955-6900, Saudi Arabia. Correspondence and requests for materials should be addressed to K.Y. (email: kyamane@gifu-u.ac.jp)
DOI番号:10.1038/s41598-019-49667-z
論文公開URL:www.nature.com/articles/s41598-019-49667-z
【研究支援】
本研究はJSPS科研費 JP15K07289 およびJP24510325の支援を受けました。
【用語解説】
1) PCR [Polymerase Chain Reaction]
ごく微量なDNA検体を好熱菌に由来のDNAポリメラーゼという特殊な酵素によって大量に増幅させる方法。発明者はノーベル賞を受賞している。
2) Multiplex PCR [Multiplex PCR]
複数のプライマー配列(特定の断片が得られるように設計された短い配列)の組み合わせを用いた反応。今回、この方法を用いることで、常にコントロールバンドが得られ、バンド無しを実験エラーと区別することができる。
3) PCR-RFLP [PCR- Restriction Fragment Length Polymorphism]
制限酵素という、特定の場所を切断する酵素で切断されたDNA断片の長さを比較することにより多型を検出する方法。
【その他のワサビ関連情報】
2019年10月24日に大阪市中央卸売市場で「第34回全国わさび品評会」が開催されます。全国から優秀なワサビが集まり、市場関係者や農業試験場職員、大学関係者として唯一岐阜大学の山根准教授が審査員として参加します。
本研究成果は,英国の国際誌「Scientific Reports」(電子版)2019年10月7日付(日本時間午後6時)に掲載されました。
【発表のポイント】
| 日本のワサビ属植物は、第四期の氷河期時代に大陸と日本列島が陸続きになった際に大陸から渡ってきた |
・ 日本のワサビ属植物2種7系統の葉緑体全ゲノムを世界で初めて解読した
・ 日本固有種であると考えられていたユリワサビの中には、大陸のワサビ属植物に近縁な系統が含まれていた
・ 日本固有と考えられるワサビ属植物系統群は約130万年前に分岐したことがわかった
・ 栽培ワサビのルーツに北方系野生ワサビはおそらくは関与していない
・ 栽培ワサビとなった野生ワサビは日本固有種であり、氷河期時代に日本海側気候に適応した日本海要素植物であることがわかった
・ 葉緑体全ゲノム配列約15万塩基対の比較解析から、日本の主要なワサビ3品種を区別するDNAマーカーの構築に成功した。これらは品種の特定に用いることができ、品種改良や保全活動への応用が期待できる
【概要】
日本にはワサビとユリワサビという、二種類のワサビ属植物が生育しています(写真1)。日本人が薬味として利用してきた栽培ワサビは、野生のワサビが栽培化されたものです。ところが野生ワサビのルーツに関しては全くわかっていませんでした。「日本のワサビ属植物は、もともと日本列島の形成時からいた植物なのか?」あるいは「大陸からわたってきた植物なのか?」—それすら不明でした。そこで今回、葉緑体の全ゲノム配列を用いて、大陸の近縁なワサビ属植物との比較解析を行い、日本のワサビ属植物がどのように大陸から移動し、分布を広げたのかを調べました。その結果、岐阜県に自生するユリワサビが、他の日本のユリワサビよりも、日本から遠く離れた中国雲南省に自生するワサビ属植物と遺伝的に近縁であることがわかったのです。ユリワサビと考えられていた一部の系統は、日本固有のユリワサビとは異なる種である可能性が考えられます。また、分岐年代の推定から、この二種間は比較的最近(48万年前)に分岐したことがわかりました。つまり、この岐阜県のユリワサビは、他のユリワサビとは別の時代に、別の祖先系統から日本にわたってきたと考えられます。その他の日本のワサビ属植物は、分岐年代推定の結果、少なくとも約130万年前に大陸のワサビ属植物とは分化したことが明らかになりました。ただし、この日本のワサビ属植物のうち北海道のワサビは、本州および九州のワサビ属植物とは遺伝的な分化の程度が大きかったことから、少なくとも現在のワサビ品種は、北方系のワサビ属植物はルーツでないと考えられます。
以上の結果を図にまとめたものが図1になります。日本のワサビ属植物は、第四紀の氷河期時代の氷期に、陸続きになった際に大陸からわたってきた祖先に由来し、しかもこうした大陸からの移動は複数回あった可能性が高いこともわかりました。現在、野生ワサビは主に日本海側の多雪地帯に分布しています。野生ワサビの祖先種が生育していた大陸とは環境が大きく異なっていたことが予想されることから、野生ワサビは、氷河期時代に列島に渡ってきた後、日本の多雪地帯に適応し、ブナなどに代表される「日本海要素植物」として成立したと考えられます。つまり、野生ワサビは、氷河期時代に日本の環境に適応して成立した日本固有種であることが、あらためて示された結果となりました。
本研究ではさらに、葉緑体全ゲノム配列を用いて主要3品種を区別するDNAマーカーを構築しました。本マーカーを用いれば、品種改良や保全集団の選定など、幅広い分野への活用が期待できます。
図1 葉緑体全ゲノム情報をもとに得られた系統樹から推定した第四紀氷河期と間氷期におけるワサビ属植物の分布と移動の模式図。日本列島には少なくとも3系統のワサビ属植物が存在することがわかる。このうち緑色で示した現在の日本固有のワサビ、ユリワサビ系統は、繰り返される氷河期と間氷期に分布が移動するなかで、現在の地理的分布域を形成した。
【研究内容】
ワサビは学名をEutrema japonicum (Miq.) Koidz. とし、日本固有種と考えられてきました。日本にはワサビとは別に、ユリワサビ[E. tenue (Miq.) Makino]という野生植物も自生しています。ユリワサビにはワサビのような立派な根茎は無く、ゆり根のような鱗片状根茎ができます(写真1)。二種は、それぞれ典型的な形態をしている個体どうしを見比べると、植物体の大きさも、花の大きさ、形態、根茎の形など、かなり違いがみられるため、両者の区別は簡単です(写真2)。また、生育している場所も、ワサビに比べてユリワサビは標高が低く、水際というよりは渓流沿いの湿った斜面に自生するなどの違いがあります。
①
③
④
⑤
⑥
写真1 野生ワサビ(①~③)とユリワサビ(④~⑥)の形態写真。上段:地上部、中段:花序、下段:根茎。野生ワサビの根茎は、栽培ワサビほど肥大しない。
一方、ワサビ属植物[Eutrema]は主に東アジアに分布するアブラナ科植物で、26種知られています。本属で栽培化された植物はワサビだけです。このうち日本のワサビ属植物に形態学的にも最も近い植物がシャンユサイ[E. yunnanense]とよばれる野生種で、主に中国雲南省に自生しています。
ワサビは、和食に欠かせない食材として日本における重要な栽培植物ですが、遺伝や進化の研究は行われてきませんでした。近年は「ゲノム」とよばれる生命の維持に必要な最小限の単位を意味する遺伝情報が様々な生物で解読されニュースになっています。ワサビでは、葉緑体とよばれる細胞小器官に存在する小さなゲノムですら解読されていませんでした。そこで本研究では、次世代シークエンサーとサンガー法を用いて7系統の配列を解読しました(図2)。
図2. ワサビ(‘ふじだるま’)葉緑体ゲノムの物理地図(サークルマップ).
葉緑体ゲノムの塩基配列は、倍数化や組換えの影響を受けないため、進化の系譜を考察するために植物では有用なツールとして汎用されています。今回得られた葉緑体全ゲノム配列のうち、遺伝子がコードされている79全ての領域の配列を用いて、すでに解読されている大陸の近縁なワサビ属植物5系統と一緒に比較解析を行いました(図3)。その結果から、以下のことが示されました。
(1) 岐阜県のユリワサビが中国雲南省の系統(シャンユサイ)とグループを形成した。この二種間の分岐は48万年と比較的最近であった
(2) 岐阜県のユリワサビとシャンユサイのグループと、その他の日本のワサビ属植物は、約130万年前に分岐した
(3) 北海道のワサビは、他の本州のワサビ属植物からは、分岐系統学的にはなれた場所に位置した
(4) 主要栽培3品種は同一グループを形成した
(1)と(2)に関しては全くの予想外で大きな問題を含んでいました。詳しく説明しましょう。図3をみると、ユリワサビとワサビは単一グループを形成しなかった(=多系統)ことから、これからはそれぞれ独立した種とみなせない可能性があります。山根は、最近の複数の系統を用いた解析や現地情報、交配実験のデータからも、ワサビとユリワサビは、独立の種としては認めず「ワサビーユリワサビ複合種」とした方がよいとの考えに至っています。仮に、複合種を認めても、認めなかったとしても、ユリワサビのうちの1系統が、大陸のシャンユサイとグループを形成したことは大きな問題となります。この系統樹に従うと、日本固有と考えられていた種が、「側系統」となります。「側系統」とは分岐系統学で用いられる専門用語ですが、今回のような樹型が得られた場合は、日本のユリワサビもワサビも、独立の種としては認めずに、E. yunnanenseのなかに含めなければならなくなるのです。つまり、日本のワサビ属植物の固有性は否定されることになります。この点は論文でも慎重に議論しましたが、近年、大陸でもE. tenueと分類された集団が複数発見されていることから、岐阜県のユリワサビの系統は、他のユリワサビとは別種であるとすることで、残りのユリワサビとワサビの系統群(図1で示した緑色のグループ)の日本固有性が支持されました。
さらに、この結果は、日本列島に、大陸から異なるタイミングで渡ってきたワサビ属植物の存在を示すことになり、「氷河期時代に陸続きになったこと」、「その際に植物が移動したこと」が、推定した分岐年代からも裏付けられたともいえます。興味深い結果といえるでしょう。
図3 葉緑体全ゲノムのコード領域の塩基配列比較からみた最尤法系統樹。四角内は分岐年代
(百万年)。枝の下の数字はブートストラップ値(枝の信頼性を表す。100に近いほど信頼性が高い)。
グループの色は、図1と関連させた。
(百万年)。枝の下の数字はブートストラップ値(枝の信頼性を表す。100に近いほど信頼性が高い)。
グループの色は、図1と関連させた。
(3)に関しても、今回の結果は予想外でした。ワサビは標高の高い冷涼な環境に好んで生育しているため、北方系の植物の要素を持つことが予想されていました。ところが今回の結果は、その可能性は低いことを示していました。つまり、(3)で示した栽培ワサビ系統は、北海道の野生ワサビよりむしろ、九州のユリワサビの方が、系統学的に近かったのです。しかも、北海道の野生ワサビは、本州と九州の「ワサビーユリワサビ系統群」からは系統樹でも離れて位置しており、これらは、図1で示したとおり、他のグループとはルーツを別にしている可能性が示されました。この結果から、日本の栽培ワサビの祖先的な野生集団は、北方ルートで渡ってきたワサビ属植物ではなく、図1で示したようなルート(緑色)を起源とすることが示唆されたのです。この、北海道の野生ワサビに関しては、現在のところ形態上の明確な違いは見つかっていませんが、樺太地方に自生する「カラフトワサビ」である可能性があり、さらに詳しく調べる必要があります。
(4)に関連して最初に、ワサビの品種について説明しましょう。あまり知られていないかもしれませんが、ワサビにはたくさんの品種が存在します。農林水産省の品種登録サイトにも約20品種が登録されています。これまで知られている主要な品種の来歴を、文献や聞き取り調査から調べた系譜を示します(図4)。たくさんの品種が作出されてきたなかで、品種改良の母本となる基本的な系統は、主要な3系統(’だるま系’、’島根3号’、’真妻’)であることがわかります。
図4. 聞き取りや文献から調査したワサビ品種の系譜(山根、2010年を改変).
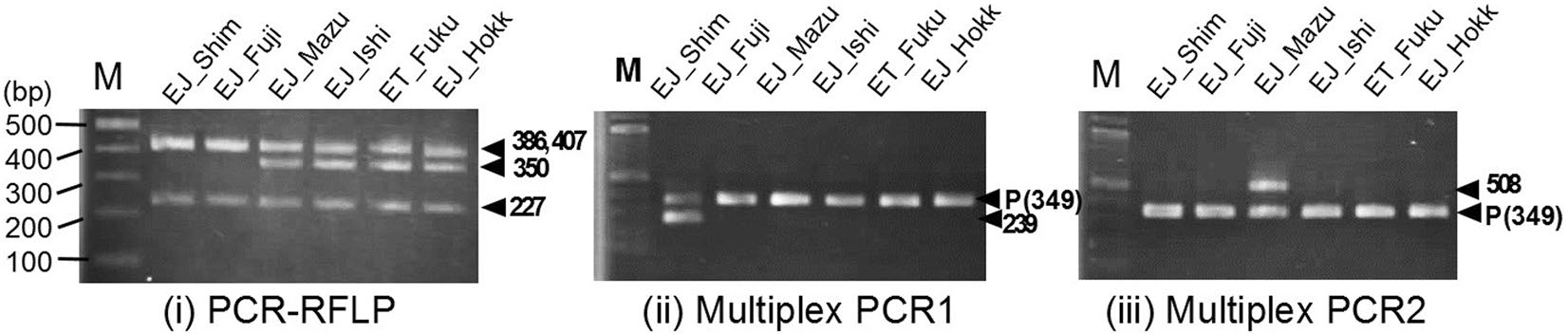
こうしたなか、品種を判別するためのDNAマーカーづくりに際し、予備的な実験を行ったところ、どうしても‘ふじだるま’と‘島根3号’を区別する多型サイトを見つけることができませんでした。つまり、両品種の母系統は遺伝的にかなり近く、区別するマーカーが見つけられなかったのです。そこで、葉緑体の全ゲノム配列を解読することで、両品種間の違いを見つけようと試みました。その結果、15万塩基対のなかで、両品種を区別するサイトは、たった2サイトしかないことがわかったのです。そこで今回、この2サイトのうちの1つを用いてDNAマーカーを構築し、両品種を区別することが可能になりました(図5)。葉緑体の全ての塩基配列を解読して初めて可能になった識別でした。
図5. ワサビ主要3品種を区別するための3種類のDNAマーカー。PCR増幅の「有る・無し」をバンド化することにより判別することができる。(i)PCR-RFLPでは、’島根3号’と’ふじだるま系’だけ350bpのバンドがみられない。(ii)MultiplexPCR1では、’島根3号’だけ239bpのバンドがみられる。そのため、(i)と(ii)を用いれば、ふじだるまと’島根3号’を区別することができる。(iii) MultiplexPCR2は、真妻だけ508bpのバンドが得られるため、’真妻’を特定するマーカーとなる。
今回構築した品種判別DNAマーカーは、あくまでも母系マーカーであり、花粉親の系統は特定できませんが、その後の多数の品種や系統を用いた結果から、現在市場に出回っている品種や、全国で商業目的に栽培されるワサビは、現在のところ例外なく今回明らかにした3タイプのDNA型しかみられないことがわかっています。このうち、’真妻’は毎年品評会で上位を占める優良品種です。’真妻’を他と区別できるマーカーが構築できたことで、品種の誤認などを回避することが可能になりました。また、’島根3号’は、’島根在来’と’半原’という病気に強い品種を自然交配により掛け合わせてできた品種であると言われてきましたが、’島根3号’の母系は、’半原’の可能性が高いことがわかりました。こうした情報を利用して、「’島根在来’さがし」の役立つツールとして用いることが可能になりました。その他「野生ワサビか栽培ワサビの逃げ出しか」の判定に、すでに今回構築したDNAマーカーは頻繁に利用されています。今後は、より詳細な品種判別を行うためにも、核DNAマーカーを開発し、より詳しく判別できるマーカーの開発が急務となっています。
【今後の展開】
本研究の目的は、大陸のワサビ属植物との比較解析を行うことで、日本のワサビ属植物のルーツを明らかにすることでした。現在は、日本全国から収集した約300系統のワサビ属植物の系統解析を行い、より詳細な分析を進めています。その結果、多くの系統を分析した結果も、今回提示した進化モデルとは矛盾がないことがわかってきました。たとえば、本研究では、中国のシャンユサイと系統学的に近い集団は岐阜県由来のユリワサビのみ供試されていましたが、その後の解析から、別の岐阜県集団や、山梨県、群馬県にも類似系統が存在することがわかったのです。ただ、これらが本当に、E. tenueとは別種であるのかどうかについては、詳細に調べる必要があります。
こうして得られた「ワサビの来た道」をふまえたうえで、現在は「日本のワサビはなぜ辛いのか」という疑問を解明するために研究を進めています。最近、海外の研究グループがワサビ属植物の辛味成分の前駆体物質を調べた論文を発表しました(Hao et al., 2016)。その結果、日本のワサビ属植物の辛味関連成分は大変ユニークであることがわかってきたのです。我々の研究室で現在も辛味関連成分を調べていますが、大変興味深いデータが得られています。
辛味関連成分が日本で独自に進化したとすると、今回の結果から、約100万年の間の第四期の氷河期時代に生じたと推測されます。日本列島に渡ってきたワサビ属植物が、氷河期時代にどのようにして独自の辛味成分を持つようになったのかを、豊富な材料を用いることで、分子レベルで明らかにしたいと考えています。
子供たちからはよく「ワサビはなぜ辛いの?」という質問を受けます。簡単な質問にみえて実は答えるのが大変難しく、本質的な問いなのだということが、研究をすすめるなかでみえてきました。現地調査や分子や化学の実験など、多角的な視野からこの本質的な問いに対する答えを見つけるべく、研究を進めています。
岐阜大学の山根研究室では、系統保存と保全活動も行っています。野生ワサビが自生する環境が年々変化し、集団の消失もみられます。将来のワサビ属植物の分布域をシミュレーション予測したところ、現在の野生ワサビの分布域は、地球温暖化により深刻な影響を受けることが示されました。日本においては、温暖化の影響として日本海側の多雪地帯の積雪量の減少が深刻になるという予測がされており、野生ワサビの自生地への影響が懸念されます。氷河期時代を生き残り、高い遺伝的多様性が維持されてきた日本のワサビ属植物は、遺伝資源として、未だ知られていない多くの可能性を秘めていることがわかってきています。日本が誇る貴重な資源を守るためにも、今後も活動を続けてゆきたいと考えています。
【論文情報】
雑誌名:Scientific Reports
タイトル:Complete chloroplast genome sequence and phylogenetic analysis of wasabi (Eutrema japonicum) and its relatives
著者:Natsuko Haga 1 , Masaaki Kobayash i2 , Nana Michiki 1 , Tomoyuki Takano 2 , Fujio Baba 3 ,
Keiko Kobayashi 1 , Hajime Ohyanagi 2,4, Jun Ohgane 2 , Kentaro Yano 2 & Kyoko Yamane1
1Gifu University, Faculty of Applied Biological Sciences, 1-1 Yanagido, Gifu City, Gifu, 501-1193, Japan. 2School of Agriculture, Meiji University, 1-1-1 Higashi-Mita, Tama-ku, Kawasaki, Kanagawa, 214-8571, Japan. 3Izu Agricultural Research Center, Shizuoka Prefectural Research Institute of Agriculture and Forestry, 3012 Inatori, Higashiizu-cho, Kamo, Shizuoka, 413-0411, Japan. 4King Abdullah University of Science and Technology (KAUST), Computational Bioscience Research Center (CBRC), Thuwal, 23955-6900, Saudi Arabia. Correspondence and requests for materials should be addressed to K.Y. (email: kyamane@gifu-u.ac.jp)
DOI番号:10.1038/s41598-019-49667-z
論文公開URL:www.nature.com/articles/s41598-019-49667-z
【研究支援】
本研究はJSPS科研費 JP15K07289 およびJP24510325の支援を受けました。
【用語解説】
1) PCR [Polymerase Chain Reaction]
ごく微量なDNA検体を好熱菌に由来のDNAポリメラーゼという特殊な酵素によって大量に増幅させる方法。発明者はノーベル賞を受賞している。
2) Multiplex PCR [Multiplex PCR]
複数のプライマー配列(特定の断片が得られるように設計された短い配列)の組み合わせを用いた反応。今回、この方法を用いることで、常にコントロールバンドが得られ、バンド無しを実験エラーと区別することができる。
3) PCR-RFLP [PCR- Restriction Fragment Length Polymorphism]
制限酵素という、特定の場所を切断する酵素で切断されたDNA断片の長さを比較することにより多型を検出する方法。
【その他のワサビ関連情報】
2019年10月24日に大阪市中央卸売市場で「第34回全国わさび品評会」が開催されます。全国から優秀なワサビが集まり、市場関係者や農業試験場職員、大学関係者として唯一岐阜大学の山根准教授が審査員として参加します。