

明治大学の瀬戸義哉准教授は静岡大学の竹内純准教授、中村彰彦教授と共同で、高温ストレス下での植物の発芽制御に関わるKAI2(KARRIKIN INSENSITIVE 2)注1というタンパク質がどのような分子構造を持つ物質(リガンド)と結びつくことで生理応答が起こるのかを明らかにしました。
地球温暖化により、高温ストレスが原因で植物の発芽が妨げられることは、農作物の収穫量に大きな影響を与える深刻な問題です。最近の研究で、植物ホルモンであるストリゴラクトン注2の受容体(D14)の仲間であるKAI2が、高温環境での植物の休眠や発芽の調節に関与していることが報告されています。しかし、KAI2と結合する植物内生リガンド注3(KL)はまだ同定されておらず、そのメカニズムもよく分かっていません。
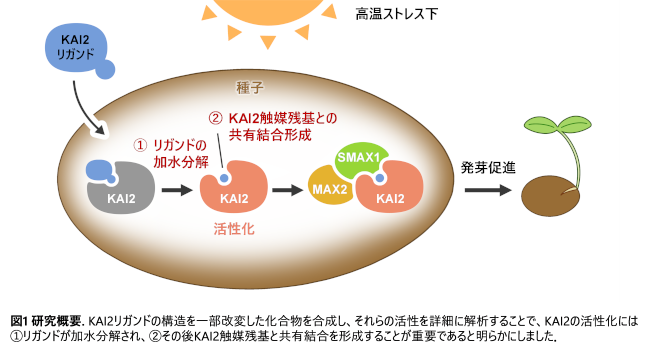
そこで本研究では、KAI2のリガンドがどのような分子構造を持っている必要があるのか、またKAI2がどのように活性化されるのかを解明しました。 具体的には、KAI2と結合することが知られているdMGer注4という物質(KAI2アゴニスト注5)の構造を改変して、KAI2と結合するが、KAI2によって加水分解されないような構造としたdMGerアナログ(類似物質)を設計しました。このアナログを使って、KAI2との結合活性や植物への効果を詳しく調べました。解析の結果、KAI2を活性化するためには、リガンドがKAI2と結合するだけでは不十分であり、リガンドのブテノライド環注6が加水分解され、その後KAI2の触媒残基注7と共有結合を形成することが重要であることが分かりました(図1)。
本成果は、10年以上発見されていなかったKLの構造的特徴に関する新しい知見を提供し、KLの探索研究を大きく前進させるものと期待されます。また、植物の高温発芽阻害の解決にKAI2経路を利用した新規農薬ターゲットを創り出せる可能性もあります。この研究成果は、2025年2月20日に国際学術誌「Proceedings of the National Academy of Sciences(PNAS)」のオンライン版で公開されました。
本成果は、以下の事業・研究領域・研究課題によって得られました。
戦略的創造研究推進事業ACT-X
研究領域:「環境とバイオテクノロジー」
(研究総括:野村 暢彦 筑波大学 生命環境系 教授)
研究課題名:「高温ストレスによる発芽阻害メカニズムの解明」(課題番号:JPMJAX21BE)
研究代表者:(竹内 純 静岡大学 准教授)
研究期間:令和3年10月~令和6年3月
研究の背景と経緯
農作物の収量が不安定になる主な理由の一つが気候変動です。全体の28~34%が気候変動の影響を受け、そのうち2~10%が高温日数の割合が増えることが原因と言われています。そのため、地球温暖化による高温障害に対応できる農業技術の開発が急務となっています。
高温障害の具体的例として、水稲栽培では米粒が白濁する白未熟粒の発生、果実栽培では着色不足、様々な農作物での発芽不良などがあります。本研究グループでは特に発芽不良(高温発芽阻害)に注目しています。高温発芽阻害は、葉茎菜類、果菜類および根菜類といった多くの農作物・植物種で生じる現象で、例えば、ホウレンソウやキャベツなど、あるいは植物研究のモデル植物として利用されるシロイヌナズナにおいては、気温が30℃以上になると発芽率が極端に低下します。この要因は主に、高温ストレスを受けた植物の体内でアブシシン酸(ABA)注8が増加し、これによって休眠が誘導されるためだと考えられています。一方で、発芽にはジベレリンやストリゴラクトン(SL)といった他の植物ホルモンも関与することが以前から知られています。また最近の研究で、SL受容体DWARF14(D14)のパラログ注9であるKARRIKIN INSENSITIVE 2(KAI2)というタンパク質が高温ストレス下での発芽制御(高温発芽阻害の緩和)に関与することが報告されています。
生体での情報伝達において、リガンドと受容体の役割は大変重要です。細胞膜などに存在する受容体(タンパク質から成る)は、情報伝達物質であるリガンドが結合すると細胞内に情報を伝えます。このとき、リガンドと受容体は1対1で対応することから、リガンドが鍵、受容体が鍵穴に例えられます。KAI2は、山火事などで植物が燃焼した際の煙に含まれる発芽刺激物質(カリキン)と結合するタンパク質として2012年に同定されました。それから10年以上経った現在でも、KAI2と結合するKLは見つかっていません。これまでの研究により、KAI2はブテノライド化合物(例えば、人工SLであるGR24ent-5DSやdMGer)と結合することで活性化することが分かっていましたが、その作用メカニズムの詳細は明らかになっていませんでした。
研究の内容
本研究では、KAI2アゴニストとして必要な分子構造やKAI2の活性化機構を明らかにすることを目的に、ブテノライド環部分を構造改変した新しいKAI2リガンドを設計・合成しました。KAI2は、α/β-加水分解酵素ファミリー注10に属するタンパク質です。特徴的なのは、受容体でありながら触媒三つ組残基(Ser-His-Asp)が保存されており、GR24ent-5DSやdMGerを加水分解する能力を持つ点です。本研究では、KAI2がリガンドを加水分解し、その後の共有結合形成がシグナル伝達にどのように関与するのかを明らかにすることを目指しました。
KAI2アゴニストであるdMGerをリード化合物として、dMGerの2-フラノン環をシクロペンテノン環に置換した1'-carba-dMGerと、フェノールエーテル結合を炭素-炭素結合に置換した6'-carba-dMGerを合成しました(図2)。これにより、1'-carba-dMGerは加水分解されず、6'-carba-dMGerは仮に加水分解されてもKAI2と共有結合しない分子構造にしました。いずれのdMGerアナログも、鏡像異性体注11を分割した後、各種試験に用いました。以降は、鏡像異性体間で活性の強かった(+)-体の結果について示しています。

(+)-1'-Carba-dMGerと(+)-6'-carba-dMGerは、いずれもin vitroの実験でKAI2と結合し、(+)-dMGerと同様に、KAI2の熱安定性を低下させました(図3A及びB)。しかし、(+)-dMGerと異なり両化合物は高温ストレスによるシロイヌナズナ種子の発芽阻害を緩和せず(図3C)、赤色光下での胚軸伸長阻害活性も(+)-dMGerのそれと比較して非常に弱いものでした(図3D)。また酵母ツーハイブリッド法注12においても、(+)-dMGerはKAI2と下流のシグナル制御因子であるSUPPRESSOR OF MAX2_1(SMAX1)との相互作用を誘導した一方、(+)-1'-carba-dMGerと(+)-6'-carba-dMGerはこの相互作用を誘導しませんでした。さらにX線結晶構造解析により、(+)-6'-carba-dMGerは加水分解されることなく、図2で示した分子構造のままKAI2の触媒ポケットに結合することが分かりました(図4)。
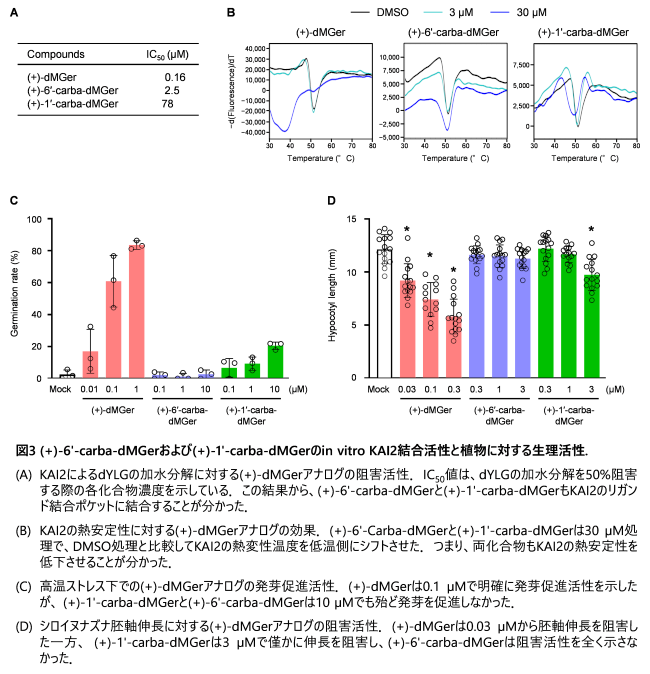
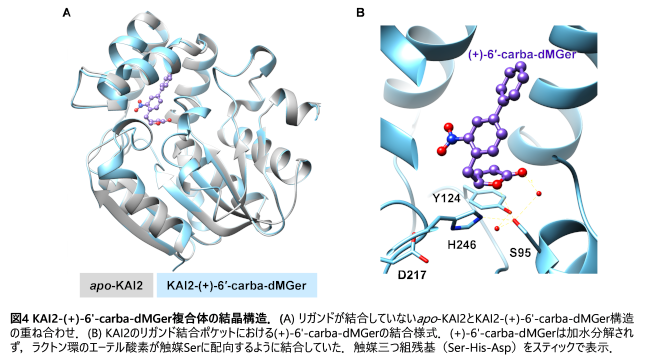
以上の結果から、KAI2シグナル伝達の活性化にはリガンドが受容体に結合するだけでは不十分であり、ブテノライド環部分の加水分解とそれに続くKAI2触媒残基との間の共有結合形成が重要な役割を担っていることを明らかにしました。従って、KLも同様に、KAI2によって加水分解・切断されるブテノライド環を有していると推測されます。
今後の展開
本成果は、10年以上にわたって未発見なKAI2の植物内生リガンドの構造的特徴に関する新たな知見を提供するものであり、新たな植物ホルモン候補であるKLの同定研究に繋がるものだと期待されます。KLが同定されれば、植物の高温ストレス応答に対するKAI2の生理機能・役割を明らかにすることができ、高温発芽阻害の制御にKAI2経路という新たな視点(新規農薬ターゲット)を創り出せる可能性があります。また本研究により得られた知見から、KAI2を人工的に制御するアゴニスト(活性化剤)やアンタゴニスト(阻害剤)の合理的な設計・合成が可能となり、KAI2を標的とした植物成長調整剤注13(環境ストレス耐性付与剤)の創出へと繋がることも期待されます。
用語解説
- 注1 KARRIKIN INSENSITIVE 2(KAI2)
山火事などで植物が燃焼した際の煙に含まれる発芽刺激物質(karrikin)と結合するタンパク質として2012年に同定された。しかし、未だ植物内生リガンド(KAI2 ligand, KL)が同定されていないオーファン受容体(オーファンとは孤児の意。特異的に結合するリガンドが同定されていない受容体)。
- 注2 ストリゴラクトン(SL)
地上部において枝分かれ・分げつを抑制する植物ホルモンである一方、根圏においては共生・寄生の化学シグナルとして作用する生理活性物質。ABAと同様に、カロテノイドの酸化開裂によって生合成される。
- 注3 植物内生リガンド
植物が生合成し、特定のタンパク質(受容体)と特異的に結合して生理活性を誘導する分子。KAI2に関しては、外因性のリガンドとしてkarrikinや人工SLが報告されているが、内生リガンド(KAI2 ligand, KL)は未だ発見されていない。
- 注4 dMGer
2023年に明治大学の瀬戸らによって開発されたKAI2選択的アゴニスト。KAI2は活性化する一方で、そのパラログであるストリゴラクトン受容体に対してはアゴニスト活性を示さない化合物。
- 注5 アゴニスト
受容体に特異的に結合し、受容体の機能を活性化することで細胞内情報伝達系を作動させる物質。KAI2にアゴニストが結合すると、KAI2の構造が不安定化して、パートナータンパク質との相互作用を介してシグナル伝達が誘導される。
- 注6 ブテノライド
ラクトンの一つで、炭素数4の複素環式化合物。最も単純なブテノライドは2-フラノンであり、天然に多く存在し、ストリゴラクトンも分子内にこの構造を含んでいる。
- 注7 触媒残基
タンパク質(酵素)が反応を触媒する際に必須となるアミノ酸残基。基質への求核攻撃を行うセリン(Ser)と、セリンの求核性を高める役割を果たすヒスチジン(His)とアスパラギン酸(Asp)の三つのアミノ酸残基がこれに相当する。KAI2においては、リガンドへの求核反応によってリガンドと触媒残基が共有結合を形成する。
- 注8 アブシシン酸(ABA)
植物ホルモン(植物が自ら生産し、微量で生体反応を制御する化合物の総称)の一つで、種子成熟・休眠維持において中心的な役割を担っている生理活性物質。乾燥ストレスを受けた植物ではABA生合成が活性化され、気孔閉鎖が誘導されることで葉からの蒸散量を抑制する。
- 注9 パラログ
進化の過程で遺伝子重複によって生じた、異なる機能を持つ遺伝子。種子植物の共通祖先で起こったKAI2の遺伝子重複によって、ストリゴラクトン受容体D14遺伝子が生じ、分化したと考えられている。
- 注10 α/β-加水分解酵素
基本骨格はβ-シートと数本のα-ヘリックス(α-ヘリックスとβ-シートはともに、アミノ酸が二次元に積み上がったタンパク質の立体構造。前者はらせん、後者は平面である)から構成されており、活性中心に触媒トライアドと呼ばれるSer-His-Asp残基を有している加水分解酵素。活性中心のセリンの水酸基が基質に求核攻撃することで触媒反応が進行する。
- 注11 鏡像異性体
立体異性体のうち、不斉炭素原子をもち、実物と鏡に映った像(鏡像)のように、原子団の位置関係が異なる異性体のこと。光学異性体ともよばれ、偏光面を回転させる旋光性の方向によって(+)-体と(−)-体が存在する。
- 注12 酵母ツーハイブリッド法
酵母内に導入したレポーター遺伝子の転写活性を指標に、タンパク質間相互作用を調べることができる手法。KAI2アゴニストは、KAI2とSMAX1の相互作用を誘導するため、本手法を用いてKAI2アゴニスト活性の有無や強弱を評価することができる。
- 注13 植物成長調整剤
農作物などの成長や発育をコントロールして、品質を高めたり、収量を増加させたり、不良条件(環境ストレス下)でも収量を安定させたり、生産上の労力を省いたりするために用いる薬剤。Plant Growth Regulator(PGR)などとも呼ばれることがある。
論文情報
タイトル
Structural requirements of KAI2 ligands for activation of signal transduction
著者名
Rito Kushihara, Akihiko Nakamura, Katsuki Takegami, Yoshiya Seto, Yusuke Kato, Hideo Dohra, Toshiyuki Ohnishi, Yasushi Todoroki, and Jun Takeuchi*
雑誌
Proceedings of the National Academy of Sciences
DOI:10.1073/pnas.2414779122
お問い合わせ先
<研究に関すること>
竹内 純(タケウチ ジュン)
静岡大学 准教授
〒422-8529 静岡県静岡市駿河区大谷836
Tel:054-238-4881
E-mail:takeuchi.jun@shizuoka.ac.jp
瀬戸 義哉(セト ヨシヤ)
明治大学 准教授
〒214-8571 神奈川県川崎市多摩区東三田1丁目1-1
E-mail:yoshiya@meiji.ac.jp
<JST事業に関すること>
原田 千夏子(ハラダ チカコ)
科学技術振興機構 戦略研究推進部 先進融合研究グループ
〒102-0076 東京都千代田区五番町7 K’s五番町
Tel:03-6380-9130 Fax:03-3222-2066
E-mail:act-x@jst.go.jp
<報道担当>
静岡大学 広報・基金課
〒422-8529 静岡県静岡市駿河区大谷836
Tel:054-238-5179 Fax:054-238-4450
E-mail:koho_all@adb.shizuoka.ac.jp
明治大学 経営企画部広報課
〒101-8301 東京都千代田区神田駿河台1-1
Tel:03-3296-4082 Fax:03-3296-4087
E-mail:koho@mics.meiji.ac.jp
科学技術振興機構 広報課
〒102-8666 東京都千代田区四番町5番地3
Tel:03-5214-8404 Fax:03-5214-8432
E-mail:jstkoho@jst.go.jp