

発表のポイント
・ シロイヌナズナの他殖性から自殖性への進化は、自家不和合性を制御する花粉遺伝子SCRに生じた逆位と転写量低下を引き起こす変異の両方が原因。
・ 高等植物の自殖と他殖を自由に制御できる可能性を開いた。
・ アブラナ科野菜の効率的な品種改良への応用が期待される。
概要
東北大学大学院生命科学研究科の渡辺正夫教授らの研究グループは、三重大学、テキサス工科大学、明治大学、東京大学、チューリッヒ大学、横浜市立大学、大阪教育大学との共同研究により、植物研究のモデル生物であるシロイヌナズナが自身の花粉で子孫(種子)を残す「自殖」へと進化した原因が花粉で機能するたった1個のSCR*2遺伝子の2種類の変異にあり、ゲノム情報と転写の二段階による変異のシナジー効果*3で制御されていることを明らかにしました。明治大学・バイオインフォマティクス研究室では、独自に整備し続けているオミックス・ビッグデータ解析基盤を活用し、大規模なDNA配列情報解析を担当しました。
本成果は、2020年9月11日、スイス科学誌「Frontiers in Plant Science」(電子版)に掲載されました。本研究は文部科学省科学研究費補助金、日本学術振興会科学研究費、スイス科学基金の支援を受けて行われました。
研究の背景
アブラナ科植物には、キャベツ、ハクサイ、ダイコンのような多くの野菜が属し、二倍体種の多くは自家不和合性を有しています。しかしながら、同じアブラナ科に属するモデル植物であるシロイヌナズナは自身の花粉で子孫を残すことができます。なぜシロイヌナズナは自殖性(自家和合性)になったのか、自家和合性への進化にはどのようなことが起きたのかと言うことは、この研究のパイオニアであるダーウィンも着目していたものの、その原因は謎でした。
研究の成果
将来への展望として、今まで不可能であったモデル生物での自家不和合性研究を可能にするとともに、アブラナ科野菜の品種改良の効率を高めるための基礎研究が飛躍的に発展することが期待できます。
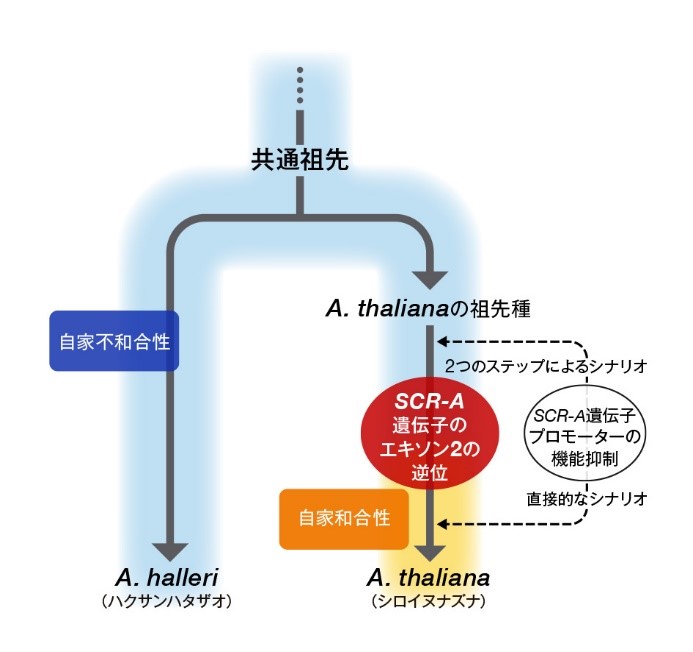
図. シロイヌナズナの自殖への進化モデル図
アブラナ科共通祖先種は自家不和合性を有しており、そこから分岐したハクサンハタザオ(A. halleri)は現在においてもそれを受け継ぎ維持している(図左側)。それに対して、シロイヌナズナ(A. thaliana)は、進化の過程においてSCRタンパク質をコードするエキソン領域と転写の制御領域にそれぞれ変異が生じ、自家不和合性を失い自殖性になったと考えられる(図右側)。
アブラナ科共通祖先種は自家不和合性を有しており、そこから分岐したハクサンハタザオ(A. halleri)は現在においてもそれを受け継ぎ維持している(図左側)。それに対して、シロイヌナズナ(A. thaliana)は、進化の過程においてSCRタンパク質をコードするエキソン領域と転写の制御領域にそれぞれ変異が生じ、自家不和合性を失い自殖性になったと考えられる(図右側)。
用語説明
*2SCR:アブラナ科自家不和合性の♂側原因タンパク質。花粉表面に付着した小型のタンパク質で、受容体である♀側原因タンパク質SRKと結合する。SP11とも呼ばれる。
*3シナジー効果:自家和合性に変異する過程において、SCR遺伝子の「エキソン2領域の逆位」と「転写を司る制御領域」が、相乗的に機能することで、自家和合性形質を安定させている。このSCR遺伝子で起きた2つの変異で生じる効果をシナジー効果と呼んでいる。
論文題目
著者: Keita Suwabe, Kaori Nagasaka, Endang Ayu Windari, Chihiro Hoshiai, Takuma Ota, Maho Takada, Ai Kitazumi, Hiromi Masuko-Suzuki, Yasuaki Kagaya, Kentaro Yano, Takashi Tsuchimatsu, Kentaro K. Shimizu, Seiji Takayama, Go Suzuki & Masao Watanabe
筆頭著者情報:諏訪部圭太、三重大学大学院生物資源学研究科
雑誌:Journal:Frontiers in Plant Science, 11: 576140
DOI: https://doi.org/10.3389/fpls.2020.576140