

概要
本成果は、2022年3月28日(現地時刻)に米国の国際学術誌「Proceedings of the National Academy of Sciences of the United States of America」にオンライン掲載されます。
1.背景
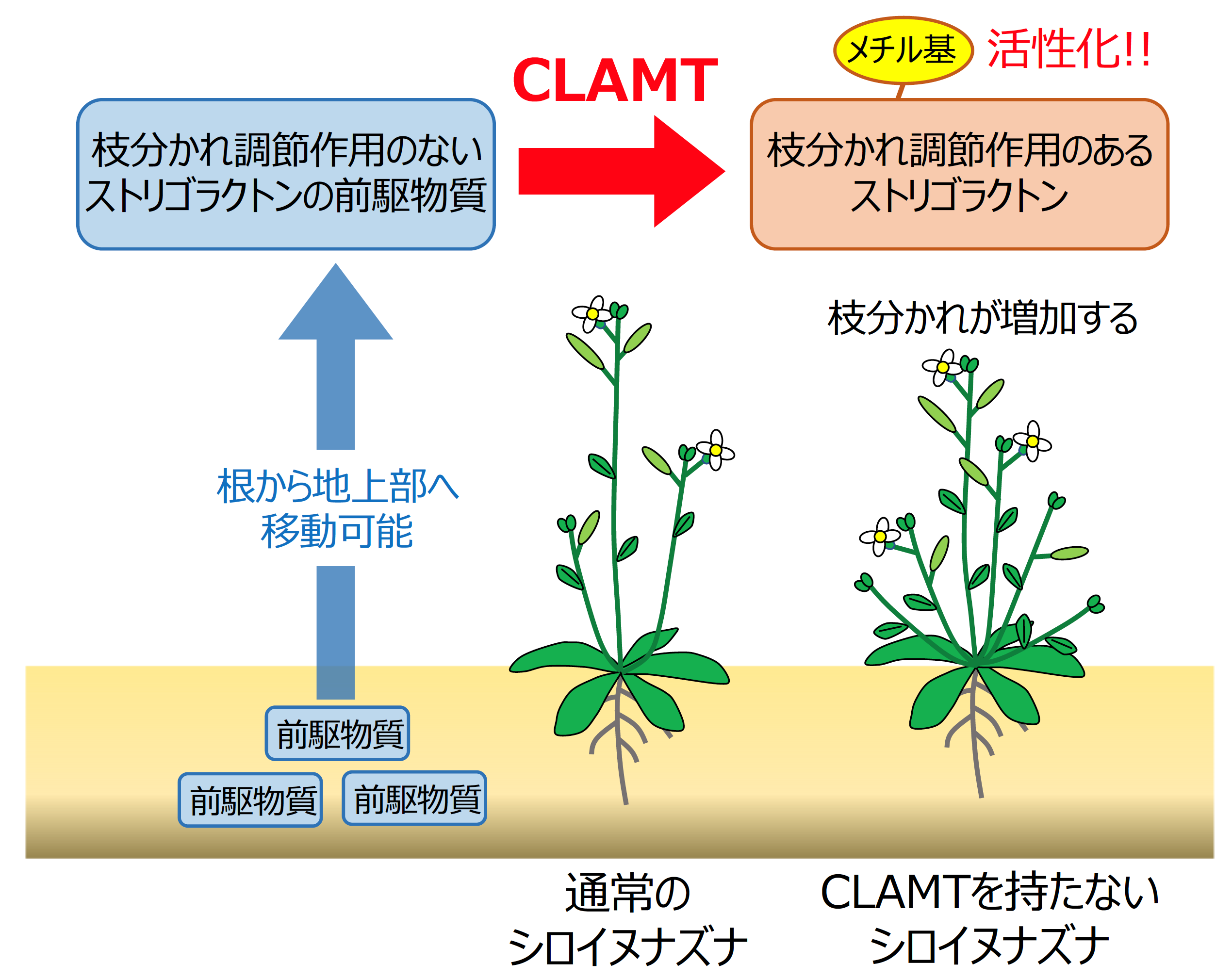
私たちの研究グループでは、これまでに数種の酵素タンパク質によって、ストリゴラクトンが植物体内でカロテノイド(β-カロテン)から、カーラクトンやカーラクトン酸と呼ばれる前駆物質を経由して作られることを明らかにしてきました(図1)。また、カーラクトン酸は未知の酵素タンパク質によって「カーラクトン酸メチル」いう物質に変換されることが予想されていました。このカーラクトン酸メチルは、植物体内でストリゴラクトンの情報を流すために必要なタンパク質(受容体タンパク質)と結合したことから、枝分かれ調節作用を持つ活性型のストリゴラクトンの1つであると考えられていました(図1)。そのため、カーラクトン酸からカーラクトン酸メチルへの変換を行う未知の酵素タンパク質を見つけ出すことは、植物体内で植物ホルモンとして働いているストリゴラクトンの形を明らかにする上で重要な研究課題となっていました。
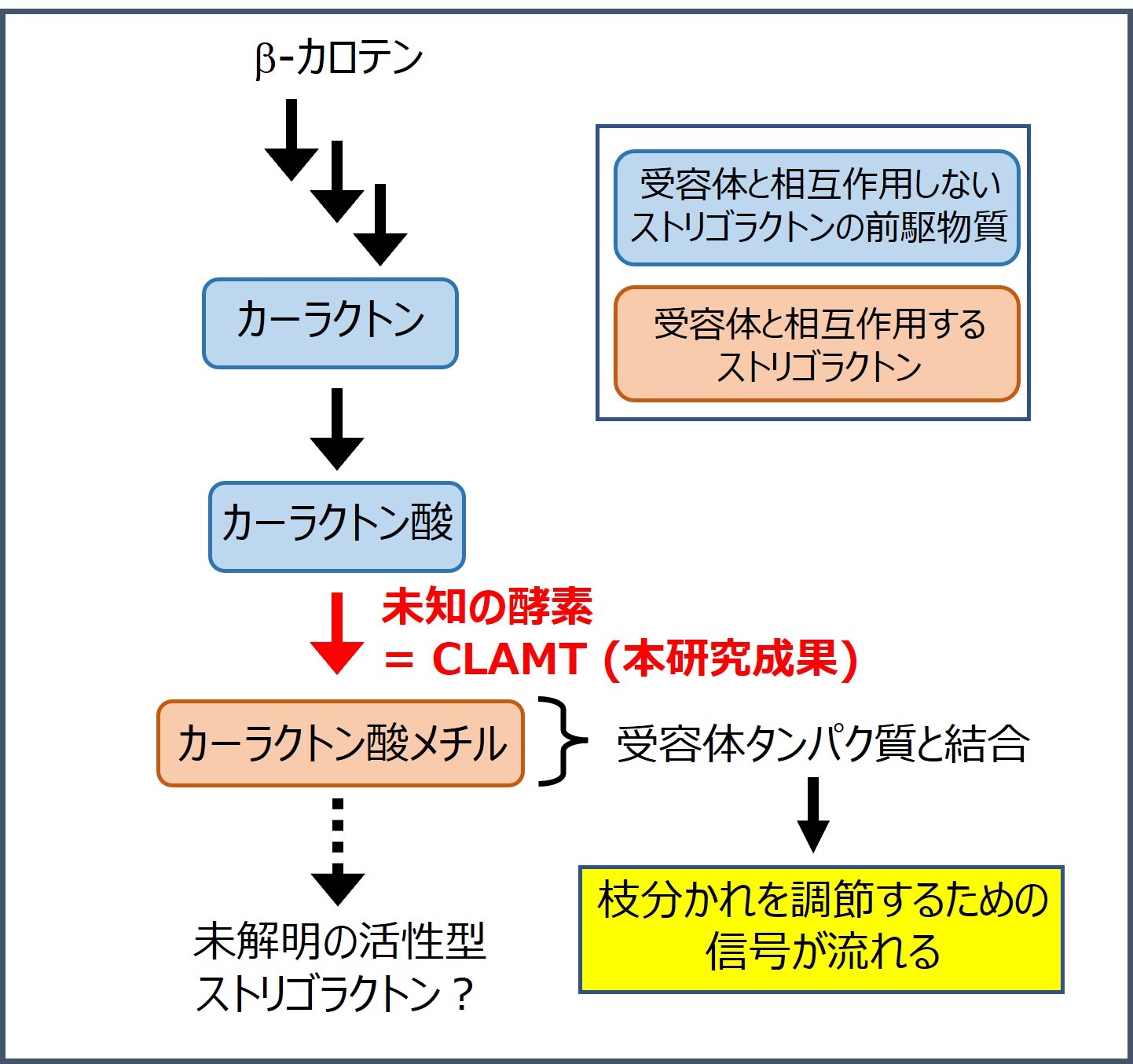
図1 植物(シロイヌナズナ)におけるストリゴラクトンの作られ方
これまでの研究から、カーラクトン酸メチルは、カーラクトンとカーラクトン酸を経由して作られると予想されていた。また、カーラクトンとカーラクトン酸は受容体タンパク質と相互作用しないが、カーラクトン酸メチルは相互作用することが示されていた。
これまでの研究から、カーラクトン酸メチルは、カーラクトンとカーラクトン酸を経由して作られると予想されていた。また、カーラクトンとカーラクトン酸は受容体タンパク質と相互作用しないが、カーラクトン酸メチルは相互作用することが示されていた。
2.研究手法・成果
また、過去に主に接木[注6]を利用して行われてきた研究により、ストリゴラクトンは根から地上部へと移動すると提唱されてきました。私たちは、CLAMT欠損変異体などを利用した接木実験によって、シロイヌナズナの体内では、植物ホルモンとして活性を持たないカーラクトンとカーラクトン酸は根から地上部へと移動可能であるものの、活性を有するカーラクトン酸メチル自身は移動しない可能性があることを明らかにしました(図3)。植物は根から地上部へと活性を持たないストリゴラクトンを移動させ、ホルモンとして作用させたい場所でCLAMTによって活性型のストリゴラクトンに変換しているという巧妙なメカニズムの存在が示唆されました。
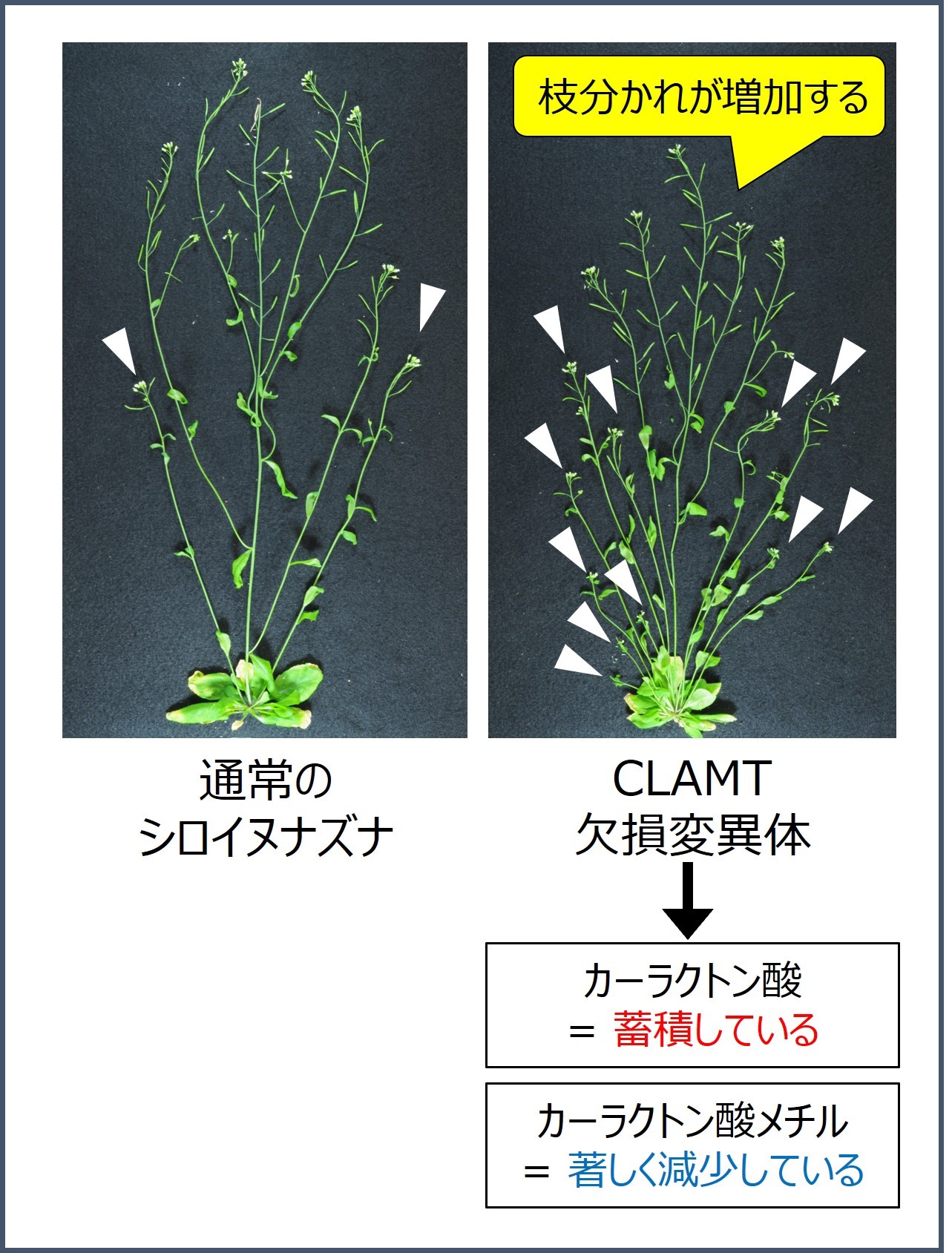
図2 CLAMT欠損変異体の解析
LC-MS/MSを用いた定量分析により、CLAMT欠損変異体ではカーラクトン酸が増加し、カーラクトン酸メチルが著しく減少していた。また、CLAMT欠損変異体は通常の植物体と比較して、枝分かれが増加したことから、CLAMTによるカーラクトン酸からカーラクトン酸メチルへの変換が、枝分かれ抑制ホルモンとしてストリゴラクトンの活性化に重要であることが示された。白い矢印は、腋芽から成長した側枝を表す。
LC-MS/MSを用いた定量分析により、CLAMT欠損変異体ではカーラクトン酸が増加し、カーラクトン酸メチルが著しく減少していた。また、CLAMT欠損変異体は通常の植物体と比較して、枝分かれが増加したことから、CLAMTによるカーラクトン酸からカーラクトン酸メチルへの変換が、枝分かれ抑制ホルモンとしてストリゴラクトンの活性化に重要であることが示された。白い矢印は、腋芽から成長した側枝を表す。
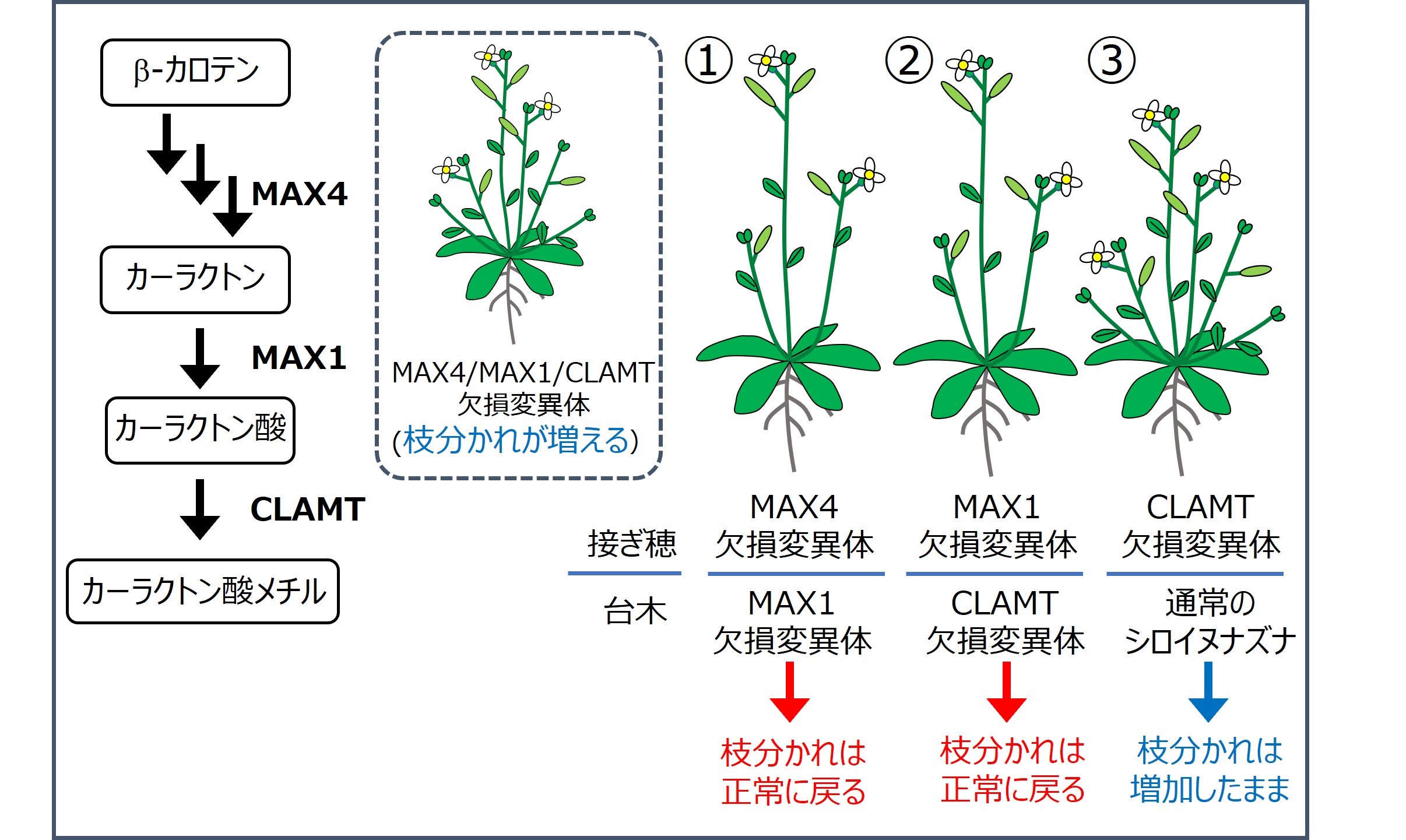
図3 接木実験結果の例
今回の研究では、接木を利用してストリゴラクトンの根から地上部への移動性も検討した。
① カーラクトン以降を作ることができないMAX1変異体の台木が、カーラクトンを作ることができないMAX4変異体の接ぎ穂で増加する枝分かれを正常に戻したので、カーラクトンが根から地上部へと移動することが示唆された。
② カーラクトン酸以降を作ることができないCLAMT変異体の台木が、カーラクトン酸を作ることができないMAX1変異体の接ぎ穂で増加する枝分かれを正常に戻したので、カーラクトン酸が根から地上部へと移動することが示唆された。
③ 通常のシロイヌナズナの台木は、カーラクトン酸メチル以降を作ることができないCLAMT変異体の接ぎ穂において増加する枝分かれを戻すことができなかったので、カーラクトン酸メチル(もしくはその代謝物)は根から地上部へと移動しないことが示唆された。(しかし、ここでは省略するが、CLAMTの下流に存在すると考えられている酵素タンパク質の欠損変異体を用いた結果では逆の結果が得られており、カーラクトン酸メチル以降の物質の移動性については今後詳細に解析する必要がある)
今回の研究では、接木を利用してストリゴラクトンの根から地上部への移動性も検討した。
① カーラクトン以降を作ることができないMAX1変異体の台木が、カーラクトンを作ることができないMAX4変異体の接ぎ穂で増加する枝分かれを正常に戻したので、カーラクトンが根から地上部へと移動することが示唆された。
② カーラクトン酸以降を作ることができないCLAMT変異体の台木が、カーラクトン酸を作ることができないMAX1変異体の接ぎ穂で増加する枝分かれを正常に戻したので、カーラクトン酸が根から地上部へと移動することが示唆された。
③ 通常のシロイヌナズナの台木は、カーラクトン酸メチル以降を作ることができないCLAMT変異体の接ぎ穂において増加する枝分かれを戻すことができなかったので、カーラクトン酸メチル(もしくはその代謝物)は根から地上部へと移動しないことが示唆された。(しかし、ここでは省略するが、CLAMTの下流に存在すると考えられている酵素タンパク質の欠損変異体を用いた結果では逆の結果が得られており、カーラクトン酸メチル以降の物質の移動性については今後詳細に解析する必要がある)
3.波及効果、今後の予定
4.研究プロジェクトについて
用語解説
[注2] 根寄生植物:ハマウツボ科植物に属するストライガやオロバンキなどの根寄生植物は、宿主植物の根から分泌されるストリゴラクトンを認識することで種子発芽が誘導される。発芽後、根寄生植物は、吸器と呼ばれる特殊な器官を使って宿主植物の根に侵入する。アフリカでは、ストライガがソルガムやトウモロコシなどの作物に寄生し、深刻な農業被害が出ており、根寄生植物の有効な駆除法の確立が望まれている。
[注3] シロイヌナズナ:アブラナ科植物に属するシロイヌナズナは、2000年に植物として初めて全ゲノム配列が解読された。世代時間が2ヶ月と短い、遺伝子組換えが容易である、変異体ラインが整備されているといった理由から、モデル植物として植物科学研究の進展に中心的な役割を果たしてきた。
[注4] 酵素タンパク質ファミリー:本研究では、「SABATHファミリー」と呼ばれるメチル化酵素群の中からCLAMTを見出した。シロイヌナズナのSABATHファミリーには24種のメチル化酵素が存在し、オーキシンやジベレリン、ジャスモン酸といった植物ホルモンをメチル化するものが含まれている。
[注5] 液体クロマトグラフ-タンデム型質量分析計(LC-MS/MS):液体クロマトグラフは化合物を分離するための装置であり、質量分析計は試料をイオン化し、分離・検出することで化合物の正確な分子量や化学構造に関する情報を得るための装置である。これら2種類の装置が接続された液体クロマトグラフ-タンデム型質量分析計(LC-MS/MS)では、2段階の質量分析(目的の化合物に由来するイオンをさらに分解して得られるイオンを測定する)ができるため、植物に極微量にしか存在しない植物ホルモンの正確な同定や定量に威力を発揮する。
[注6] 接木:2種の植物からの「台木」と「接ぎ穂」を接ぐことで、両方の性質を兼ね備えた植物体を作り出す技術。シロイヌナズナでは、切断した芽生えの胚軸(最初に伸びる茎の部分)を繋ぎ合わせることで接木を行う。これまでにシロイヌナズナやエンドウ、ペチュニアを用いた接木実験から、ストリゴラクトンは根から地上部へと移動し、地上部の枝分かれを抑制可能であることが示されてきたが、移動しているストリゴラクトンの分子実体については未解明であった。
研究者のコメント
本研究では、植物の枝分かれを調節するストリゴラクトンが作られる過程の中で、重要なステップを解明しました。しかし、過去の研究報告を照らし合わせると、カーラクトン酸メチルの他にも活性型のストリゴラクトンが存在する可能性が十分あります(図1)。まだまだ謎に包まれているストリゴラクトンの作られ方について、さらなる実験を進めることで迫っていきたいと考えています。(京都大学化学研究所 増口潔)
論文タイトルと著者
著 者:Kiyoshi Mashiguchi*, Yoshiya Seto*, Yuta Onozuka*, Sarina Suzuki, Kiyoko Takemoto, Yanting Wang, Lemeng Dong, Kei Asami, Ryota Noda, Takaya Kisugi, Naoki Kitaoka, Kohki Akiyama, Harro Bouwmeester, Shinjiro Yamaguchi†(*共筆頭著者、†責任著者)
掲 載 誌:Proceedings of the National Academy of Sciences of the United States of America
DOI:10.1073/pnas.2111565119